

MicroRNAs Modulate Signaling Pathways in Osteogenic Differentiation of Mesenchymal Stem Cells
- PMID: 33673409
- PMCID: PMC7956574
- DOI: 10.3390/ijms22052362
MicroRNAs Modulate Signaling Pathways in Osteogenic Differentiation of Mesenchymal Stem Cells
Abstract
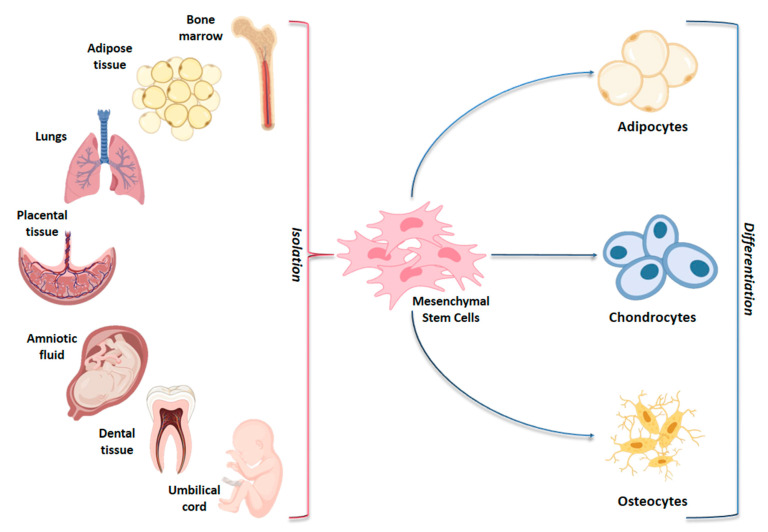
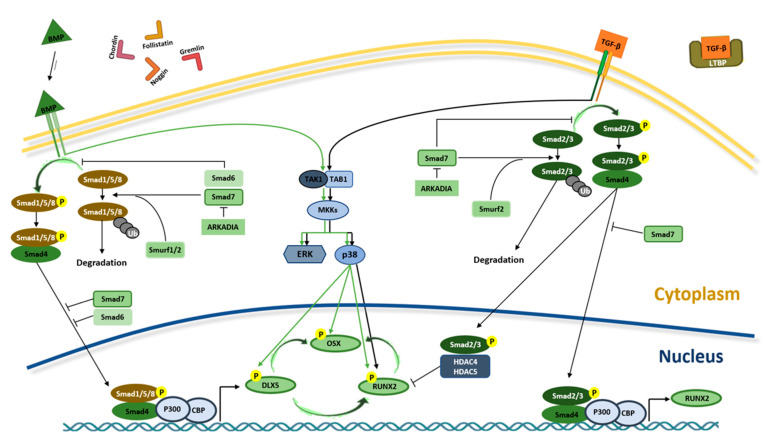
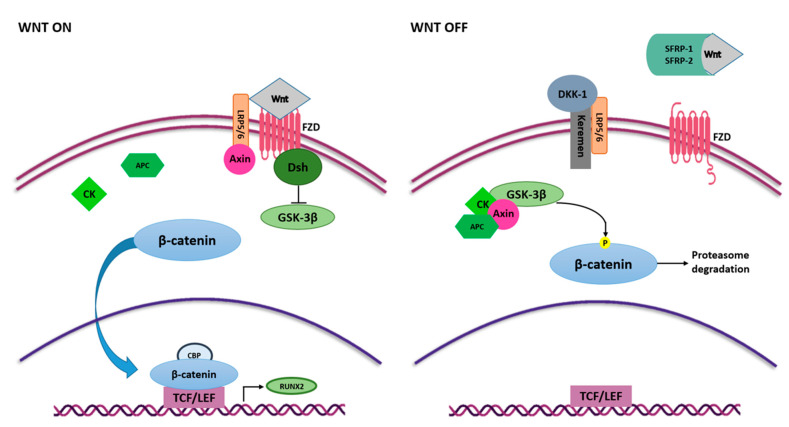
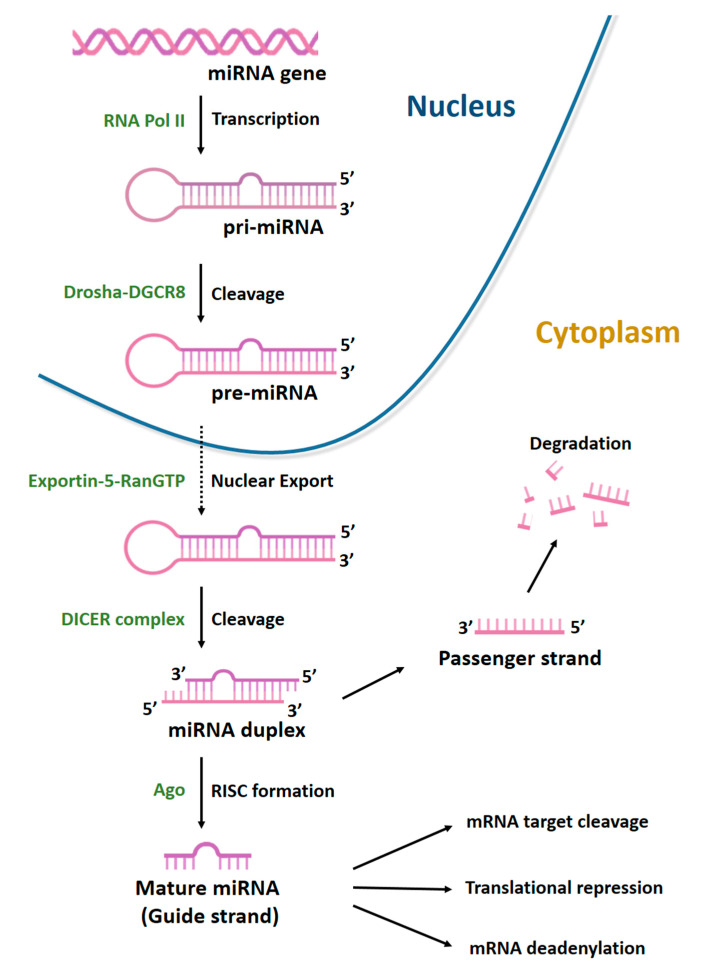
Mesenchymal stem cells (MSCs) have been identified in many adult tissues and they have been closely studied in recent years, especially in view of their potential use for treating diseases and damaged tissues and organs. MSCs are capable of self-replication and differentiation into osteoblasts and are considered an important source of cells in tissue engineering for bone regeneration. Several epigenetic factors are believed to play a role in the osteogenic differentiation of MSCs, including microRNAs (miRNAs). MiRNAs are small, single-stranded, non-coding RNAs of approximately 22 nucleotides that are able to regulate cell proliferation, differentiation and apoptosis by binding the 3' untranslated region (3'-UTR) of target mRNAs, which can be subsequently degraded or translationally silenced. MiRNAs control gene expression in osteogenic differentiation by regulating two crucial signaling cascades in osteogenesis: the transforming growth factor-beta (TGF-β)/bone morphogenic protein (BMP) and the Wingless/Int-1(Wnt)/β-catenin signaling pathways. This review provides an overview of the miRNAs involved in osteogenic differentiation and how these miRNAs could regulate the expression of target genes.
Keywords: MSCs; bone; bone regeneration; cell differentiation; mesenchymal stem cells; miRNA; miRNAome; microRNA; osteogenesis; osteogenic differentiation; stem cells.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Mazzoni E., D’Agostino A., Iaquinta M.R., Bononi I., Trevisiol L., Rotondo J.C., Patergnani S., Giorgi C., Gunson M.J., Arnett G.W., et al. Hydroxylapatite-Collagen Hybrid Scaffold Induces Human Adipose-Derived Mesenchymal Stem Cells to Osteogenic Differentiation in Vitro and Bone Regrowth in Patients. Stem Cells Transl. Med. 2020;9:377–388. doi: 10.1002/sctm.19-0170. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
