

Unique Human and Mouse β-Cell Senescence-Associated Secretory Phenotype (SASP) Reveal Conserved Signaling Pathways and Heterogeneous Factors
- PMID: 33674410
- PMCID: PMC8173799
- DOI: 10.2337/db20-0553
Unique Human and Mouse β-Cell Senescence-Associated Secretory Phenotype (SASP) Reveal Conserved Signaling Pathways and Heterogeneous Factors
Abstract
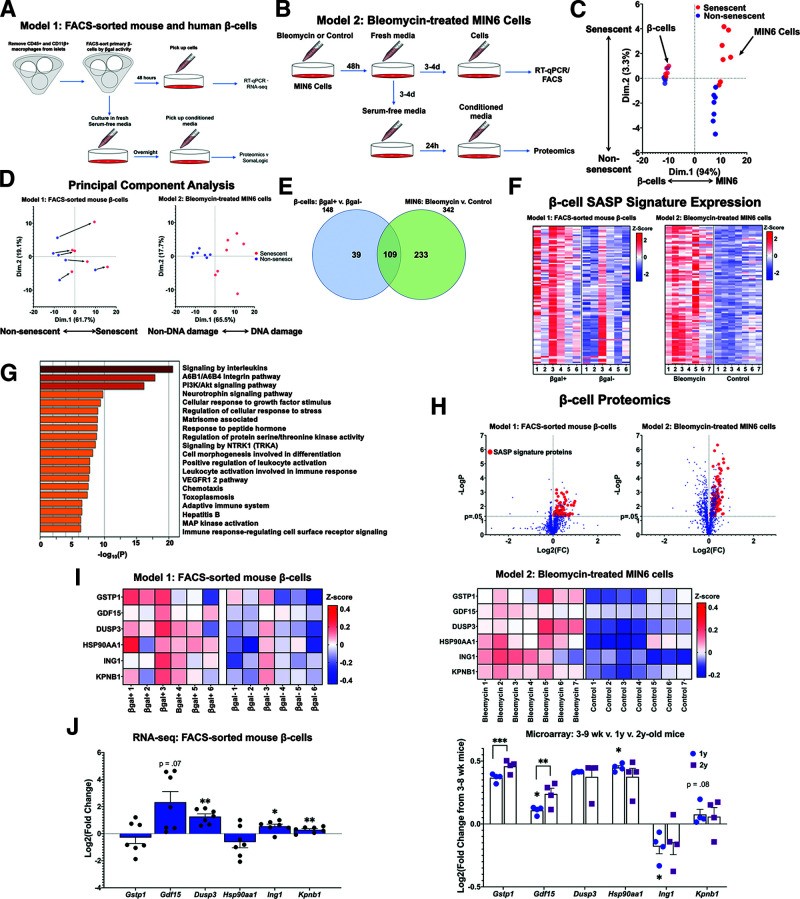
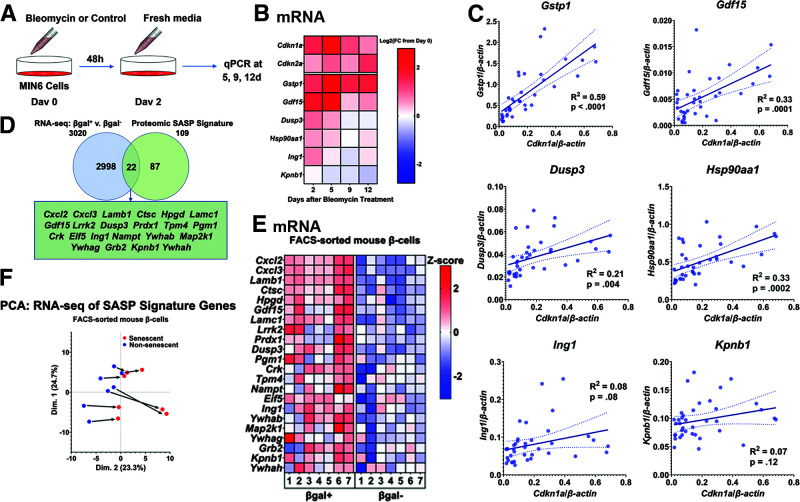
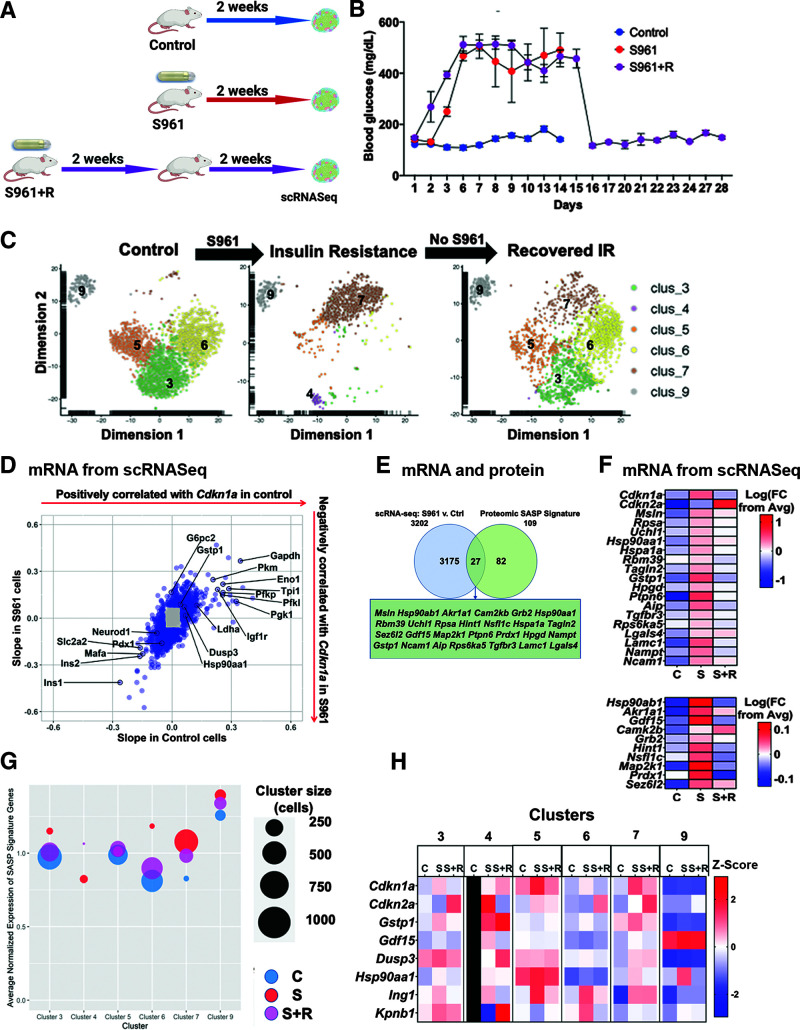
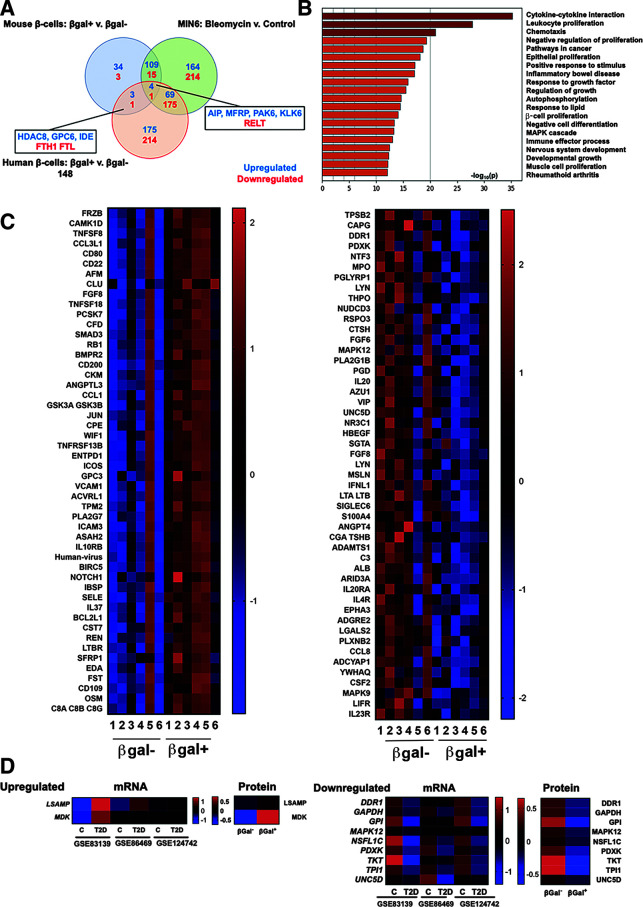
The aging of pancreatic β-cells may undermine their ability to compensate for insulin resistance, leading to the development of type 2 diabetes (T2D). Aging β-cells acquire markers of cellular senescence and develop a senescence-associated secretory phenotype (SASP) that can lead to senescence and dysfunction of neighboring cells through paracrine actions, contributing to β-cell failure. In this study, we defined the β-cell SASP signature based on unbiased proteomic analysis of conditioned media of cells obtained from mouse and human senescent β-cells and a chemically induced mouse model of DNA damage capable of inducing SASP. These experiments revealed that the β-cell SASP is enriched for factors associated with inflammation, cellular stress response, and extracellular matrix remodeling across species. Multiple SASP factors were transcriptionally upregulated in models of β-cell senescence, aging, insulin resistance, and T2D. Single-cell transcriptomic analysis of islets from an in vivo mouse model of reversible insulin resistance indicated unique and partly reversible changes in β-cell subpopulations associated with senescence. Collectively, these results demonstrate the unique secretory profile of senescent β-cells and its potential implication in health and disease.
© 2021 by the American Diabetes Association.
Figures
Similar articles
-
Cellular Senescence in Diabetes Mellitus: Distinct Senotherapeutic Strategies for Adipose Tissue and Pancreatic β Cells.Front Endocrinol (Lausanne). 2022 Mar 31;13:869414. doi: 10.3389/fendo.2022.869414. eCollection 2022. Front Endocrinol (Lausanne). 2022. PMID: 35432205 Free PMC article. Review.
-
Acceleration of β Cell Aging Determines Diabetes and Senolysis Improves Disease Outcomes.Cell Metab. 2019 Jul 2;30(1):129-142.e4. doi: 10.1016/j.cmet.2019.05.006. Epub 2019 May 30. Cell Metab. 2019. PMID: 31155496 Free PMC article.
-
Senescent myoblasts exhibit an altered exometabolome that is linked to senescence-associated secretory phenotype signaling.Am J Physiol Cell Physiol. 2025 Feb 1;328(2):C440-C451. doi: 10.1152/ajpcell.00880.2024. Epub 2024 Dec 26. Am J Physiol Cell Physiol. 2025. PMID: 39726265
-
BET Proteins Are Required for Transcriptional Activation of the Senescent Islet Cell Secretome in Type 1 Diabetes.Int J Mol Sci. 2019 Sep 26;20(19):4776. doi: 10.3390/ijms20194776. Int J Mol Sci. 2019. PMID: 31561444 Free PMC article.
-
Spatial and Temporal Control of Senescence.Trends Cell Biol. 2017 Nov;27(11):820-832. doi: 10.1016/j.tcb.2017.07.004. Epub 2017 Aug 17. Trends Cell Biol. 2017. PMID: 28822679 Review.
Cited by
-
The Importance of Intra-Islet Communication in the Function and Plasticity of the Islets of Langerhans during Health and Diabetes.Int J Mol Sci. 2024 Apr 6;25(7):4070. doi: 10.3390/ijms25074070. Int J Mol Sci. 2024. PMID: 38612880 Free PMC article. Review.
-
Endocrine-exocrine miR-503-322 drives aging-associated pancreatitis via targeting MKNK1 in acinar cells.Nat Commun. 2025 Mar 17;16(1):2613. doi: 10.1038/s41467-025-57615-x. Nat Commun. 2025. PMID: 40097383 Free PMC article.
-
The Role of Sensory Nerves in Dental Pulp Homeostasis: Histological Changes and Cellular Consequences after Sensory Denervation.Int J Mol Sci. 2024 Jan 17;25(2):1126. doi: 10.3390/ijms25021126. Int J Mol Sci. 2024. PMID: 38256202 Free PMC article.
-
Exercise activates AMPK in mouse and human pancreatic islets to decrease senescence.Nat Metab. 2024 Oct;6(10):1976-1990. doi: 10.1038/s42255-024-01130-8. Epub 2024 Sep 24. Nat Metab. 2024. PMID: 39317751 Free PMC article.
-
A narrative review on inflammaging and late-onset hypogonadism.Front Endocrinol (Lausanne). 2024 Jan 17;15:1291389. doi: 10.3389/fendo.2024.1291389. eCollection 2024. Front Endocrinol (Lausanne). 2024. PMID: 38298378 Free PMC article. Review.
References
-
- Weir GC, Bonner-Weir S.. Five stages of evolving beta-cell dysfunction during progression to diabetes. Diabetes 2004;53(Suppl. 3):S16–S21 - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
