

Role of a local transcription factor in governing cellular carbon/nitrogen homeostasis in Pseudomonas fluorescens
- PMID: 33675669
- PMCID: PMC8034625
- DOI: 10.1093/nar/gkab091
Role of a local transcription factor in governing cellular carbon/nitrogen homeostasis in Pseudomonas fluorescens
Abstract
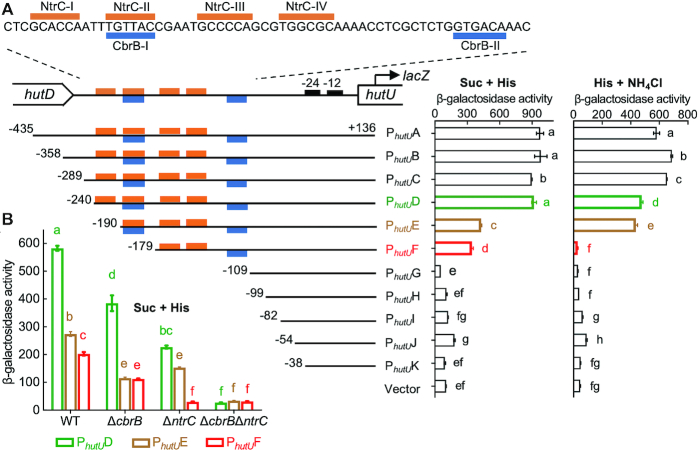
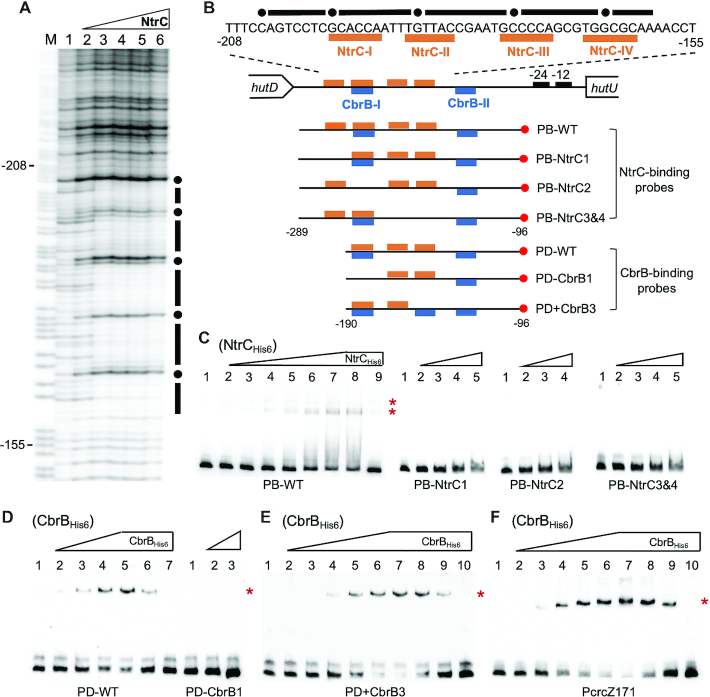
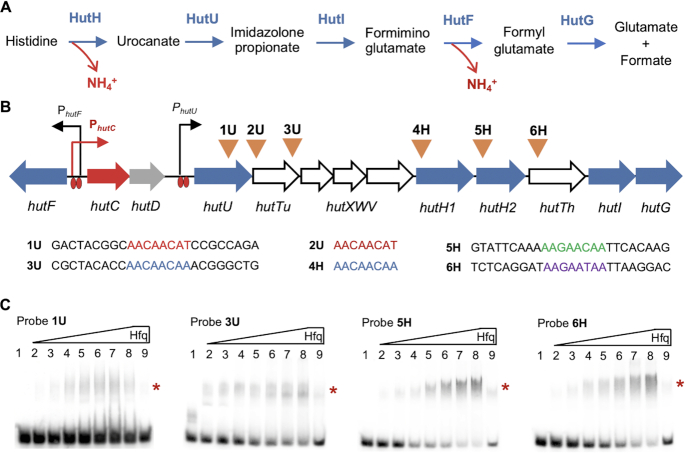
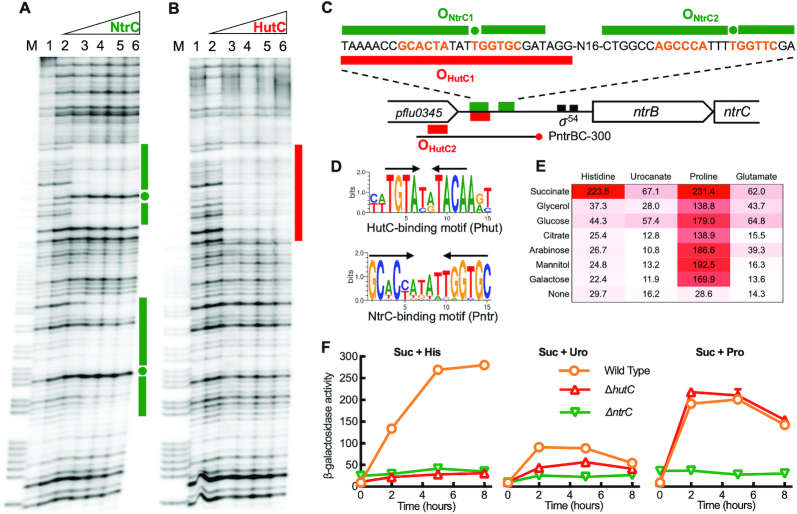
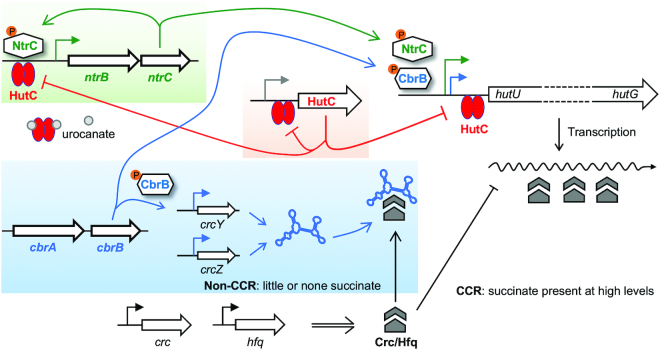
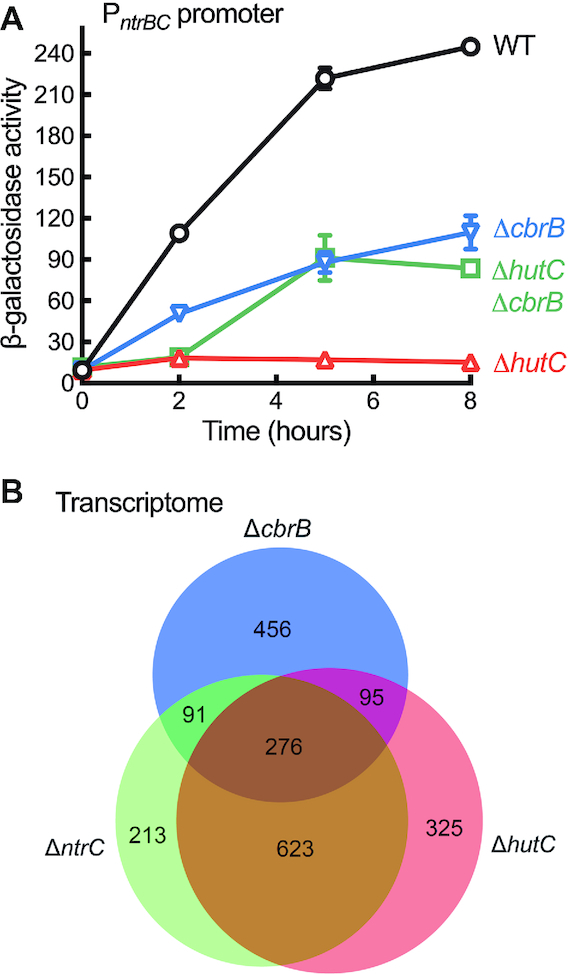
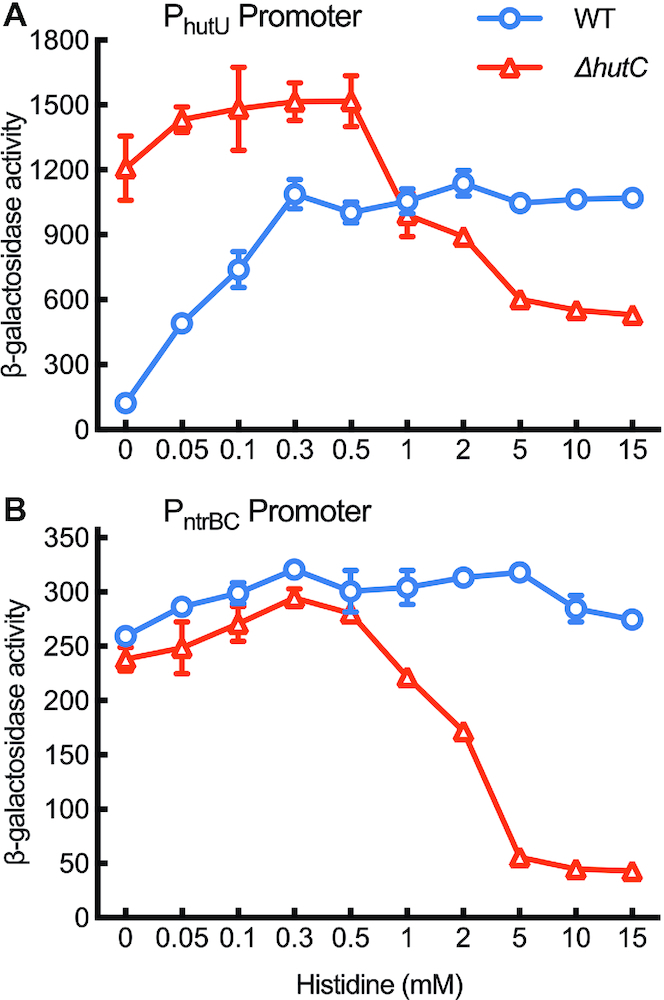
Autoactivation of two-component systems (TCSs) can increase the sensitivity to signals but inherently cause a delayed response. Here, we describe a unique negative feedback mechanism enabling the global NtrB/NtrC regulator to rapidly respond to nitrogen starvation over the course of histidine utilization (hut) in Pseudomonas fluorescens. NtrBC directly activates transcription of hut genes, but overexpression will produce excess ammonium leading to NtrBC inactivation. To prevent this from occurring, the histidine-responsive repressor HutC fine-tunes ntrBC autoactivation: HutC and NtrC bind to the same operator site in the ntrBC promoter. This newly discovered low-affinity binding site shows little sequence similarity with the consensus sequence that HutC recognizes for substrate-specific induction of hut operons. A combination of genetic and transcriptomic analysis indicated that both ntrBC and hut promoter activities cannot be stably maintained in the ΔhutC background when histidine fluctuates at high concentrations. Moreover, the global carbon regulator CbrA/CbrB is involved in directly activating hut transcription while de-repressing hut translation via the CbrAB-CrcYZ-Crc/Hfq regulatory cascade. Together, our data reveal that the local transcription factor HutC plays a crucial role in governing NtrBC to maintain carbon/nitrogen homeostasis through the complex interactions between two TCSs (NtrBC and CbrAB) at the hut promoter.
© The Author(s) 2021. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
Global Regulatory Roles of the Histidine-Responsive Transcriptional Repressor HutC in Pseudomonas fluorescens SBW25.J Bacteriol. 2020 Jun 9;202(13):e00792-19. doi: 10.1128/JB.00792-19. Print 2020 Jun 9. J Bacteriol. 2020. PMID: 32291279 Free PMC article.
-
Dual involvement of CbrAB and NtrBC in the regulation of histidine utilization in Pseudomonas fluorescens SBW25.Genetics. 2008 Jan;178(1):185-95. doi: 10.1534/genetics.107.081984. Genetics. 2008. PMID: 18202367 Free PMC article.
-
The CbrA-CbrB two-component regulatory system controls the utilization of multiple carbon and nitrogen sources in Pseudomonas aeruginosa.Mol Microbiol. 2001 May;40(4):917-31. doi: 10.1046/j.1365-2958.2001.02435.x. Mol Microbiol. 2001. PMID: 11401699
-
The Regulatory Hierarchy Following Signal Integration by the CbrAB Two-Component System: Diversity of Responses and Functions.Genes (Basel). 2022 Feb 18;13(2):375. doi: 10.3390/genes13020375. Genes (Basel). 2022. PMID: 35205417 Free PMC article. Review.
-
The role of the NAC protein in the nitrogen regulation of Klebsiella aerogenes.Mol Microbiol. 1991 Nov;5(11):2575-80. doi: 10.1111/j.1365-2958.1991.tb01965.x. Mol Microbiol. 1991. PMID: 1664020 Review.
Cited by
-
The Regulatory Functions of σ54 Factor in Phytopathogenic Bacteria.Int J Mol Sci. 2021 Nov 24;22(23):12692. doi: 10.3390/ijms222312692. Int J Mol Sci. 2021. PMID: 34884502 Free PMC article. Review.
-
Competition between Pseudomonas aeruginosa and Staphylococcus aureus is dependent on intercellular signaling and regulated by the NtrBC two-component system.Sci Rep. 2022 May 30;12(1):9027. doi: 10.1038/s41598-022-12650-2. Sci Rep. 2022. PMID: 35637237 Free PMC article.
-
Implications of carbon catabolite repression for plant-microbe interactions.Plant Commun. 2021 Dec 28;3(2):100272. doi: 10.1016/j.xplc.2021.100272. eCollection 2022 Mar 14. Plant Commun. 2021. PMID: 35529946 Free PMC article. Review.
-
NtrBC Selectively Regulates Host-Pathogen Interactions, Virulence, and Ciprofloxacin Susceptibility of Pseudomonas aeruginosa.Front Cell Infect Microbiol. 2021 Jun 24;11:694789. doi: 10.3389/fcimb.2021.694789. eCollection 2021. Front Cell Infect Microbiol. 2021. PMID: 34249781 Free PMC article.
-
Insight rifampicin-resistant (rpoB) mutation in Pseudomonas stutzeri leads to enhance the biosynthesis of secondary metabolites to survive against harsh environments.Arch Microbiol. 2022 Jun 29;204(7):437. doi: 10.1007/s00203-022-03064-9. Arch Microbiol. 2022. PMID: 35768665
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
