

Camostat mesylate inhibits SARS-CoV-2 activation by TMPRSS2-related proteases and its metabolite GBPA exerts antiviral activity
- PMID: 33676899
- PMCID: PMC7930809
- DOI: 10.1016/j.ebiom.2021.103255
Camostat mesylate inhibits SARS-CoV-2 activation by TMPRSS2-related proteases and its metabolite GBPA exerts antiviral activity
Abstract
Background: Antivirals are needed to combat the COVID-19 pandemic, which is caused by SARS-CoV-2. The clinically-proven protease inhibitor Camostat mesylate inhibits SARS-CoV-2 infection by blocking the virus-activating host cell protease TMPRSS2. However, antiviral activity of Camostat mesylate metabolites and potential viral resistance have not been analyzed. Moreover, antiviral activity of Camostat mesylate in human lung tissue remains to be demonstrated.
Methods: We used recombinant TMPRSS2, reporter particles bearing the spike protein of SARS-CoV-2 or authentic SARS-CoV-2 to assess inhibition of TMPRSS2 and viral entry, respectively, by Camostat mesylate and its metabolite GBPA.
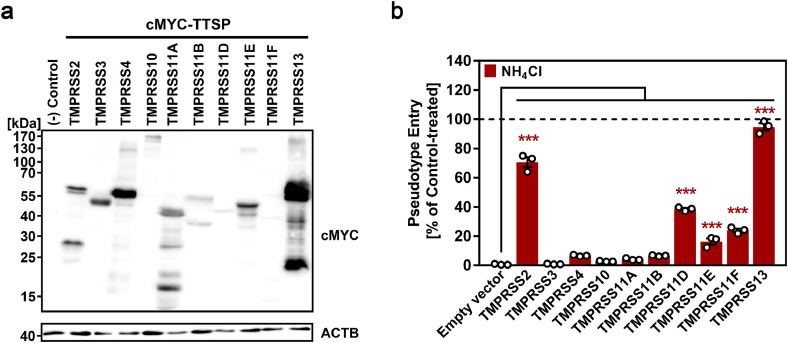
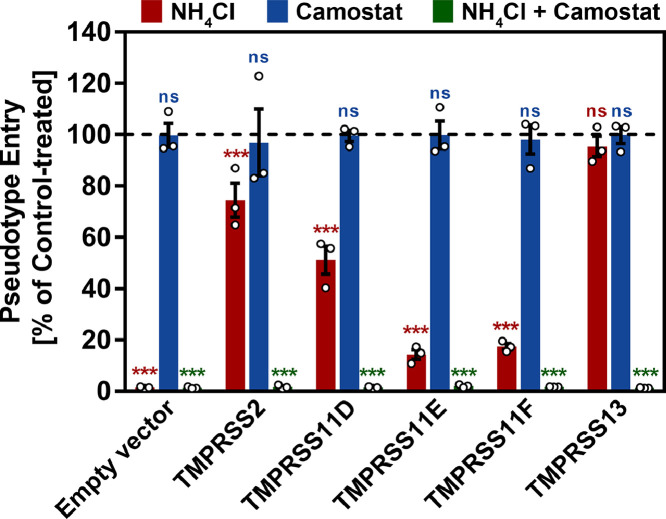
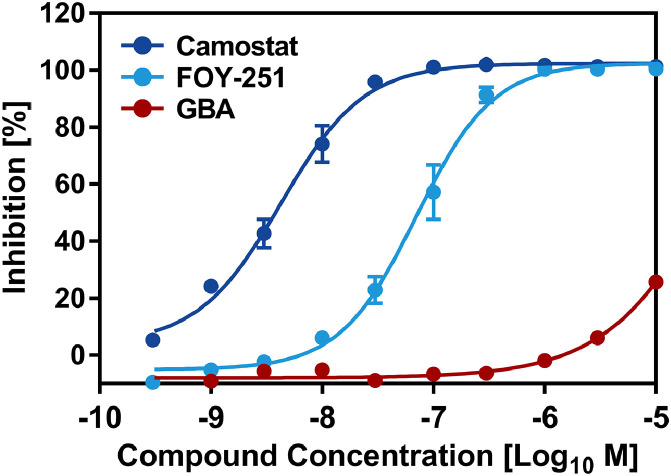
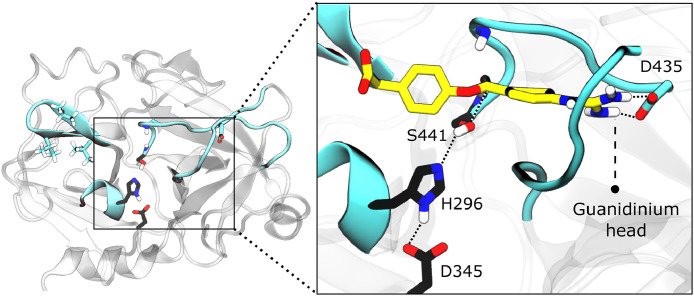
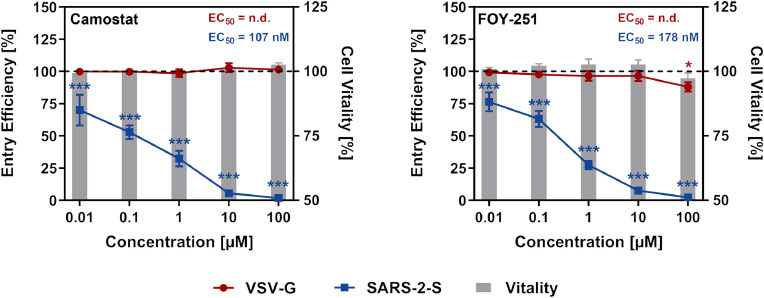
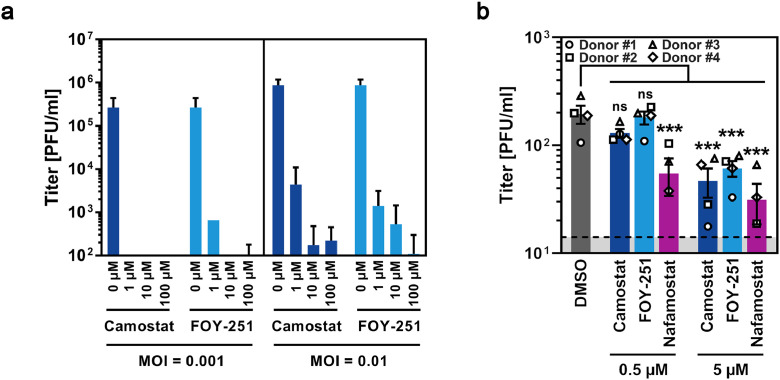
Findings: We show that several TMPRSS2-related proteases activate SARS-CoV-2 and that two, TMPRSS11D and TMPRSS13, are robustly expressed in the upper respiratory tract. However, entry mediated by these proteases was blocked by Camostat mesylate. The Camostat metabolite GBPA inhibited recombinant TMPRSS2 with reduced efficiency as compared to Camostat mesylate. In contrast, both inhibitors exhibited similar antiviral activity and this correlated with the rapid conversion of Camostat mesylate into GBPA in the presence of serum. Finally, Camostat mesylate and GBPA blocked SARS-CoV-2 spread in human lung tissue ex vivo and the related protease inhibitor Nafamostat mesylate exerted augmented antiviral activity.
Interpretation: Our results suggest that SARS-CoV-2 can use TMPRSS2 and closely related proteases for spread in the upper respiratory tract and that spread in the human lung can be blocked by Camostat mesylate and its metabolite GBPA.
Funding: NIH, Damon Runyon Foundation, ACS, NYCT, DFG, EU, Berlin Mathematics center MATH+, BMBF, Lower Saxony, Lundbeck Foundation, Novo Nordisk Foundation.
Keywords: Camostat; FOY-251; GBPA; SARS-CoV-2; TMPRSS2.
Copyright © 2021 The Authors. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of Competing Interests H.H-W., N.K., L.K.S., O.S.S., J.B.H., M.W., S.O., O.D., D.J., S.L., F.N., M.K. have nothing to disclose. T.Y., K.Y., H.M. report personal fees from Ono Pharmaceutical, during the conduct of the study. M.H. reports grants from Deutsche Forschungsgemeinschaft (DFG, German Research Foundation), during the conduct of the study. J.C.S. reports personal fees from Google, personal fees from Meliora Therapeutics, outside the submitted work. J.C.S. is an employee of Google. This work was performed outside of her affiliation with Google and used no proprietary knowledge or materials from Google. P.A. reports grants from Country of Lower Saxony, during the conduct of the study. T. H. reports grants from Deutsche Forschungsgemeinschaft (DFG) SFB/TRR 186, during the conduct of the study. L.R. reports grants from Bayer AG, outside the submitted work. A.B. reports grants from Fraunhofer DRECOR (Drug Repurposing for Corona), during the conduct of the study; other from Fraunhofer ITEM, outside the submitted work. J.M.S. reports grants from NIH, grants from New York Community Trust, grants from Damon Runyon Foundation, grants from American Cancer Society, grants from Department of Defense, during the conduct of the study; personal fees from Meliora Therapeutics, personal fees from Tyra Biosciences, personal fees from Ono Pharmaceutical, outside the submitted work. S.P. reports grants from Bundesministerium für Bildung und Forschung, grants from Deutsche Forschungsgemeinschaft, grants from Country of Lower Saxony, during the conduct of the study; other from Ono Pharmaceutical, outside the submitted work.
Figures


Update of
-
Camostat mesylate inhibits SARS-CoV-2 activation by TMPRSS2-related proteases and its metabolite GBPA exerts antiviral activity.bioRxiv [Preprint]. 2020 Aug 5:2020.08.05.237651. doi: 10.1101/2020.08.05.237651. bioRxiv. 2020. Update in: EBioMedicine. 2021 Mar;65:103255. doi: 10.1016/j.ebiom.2021.103255. PMID: 32793911 Free PMC article. Updated. Preprint.
Comment in
-
COVID19 therapeutics: Expanding the antiviral arsenal.EBioMedicine. 2021 Apr;66:103289. doi: 10.1016/j.ebiom.2021.103289. Epub 2021 Mar 19. EBioMedicine. 2021. PMID: 33752131 Free PMC article. No abstract available.
References
-
- World Health Organization (WHO) Coronavirus disease (COVID-19) Situation Report. 2020;184
-
- Beigel JH, Tomashek KM, Dodd LE. Remdesivir for the treatment of COVID-19 - preliminary report. N Engl J Med. 2020 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
