

Sex-Specific Control of Human Heart Maturation by the Progesterone Receptor
- PMID: 33682422
- PMCID: PMC8055196
- DOI: 10.1161/CIRCULATIONAHA.120.051921
Sex-Specific Control of Human Heart Maturation by the Progesterone Receptor
Abstract
Background: Despite in-depth knowledge of the molecular mechanisms controlling embryonic heart development, little is known about the signals governing postnatal maturation of the human heart.
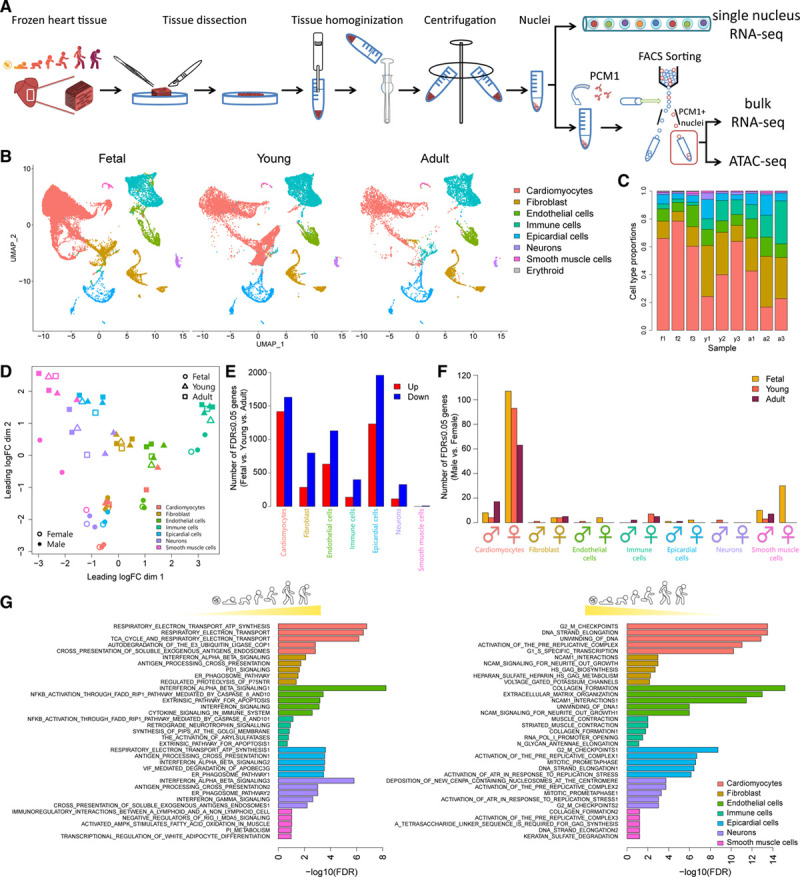
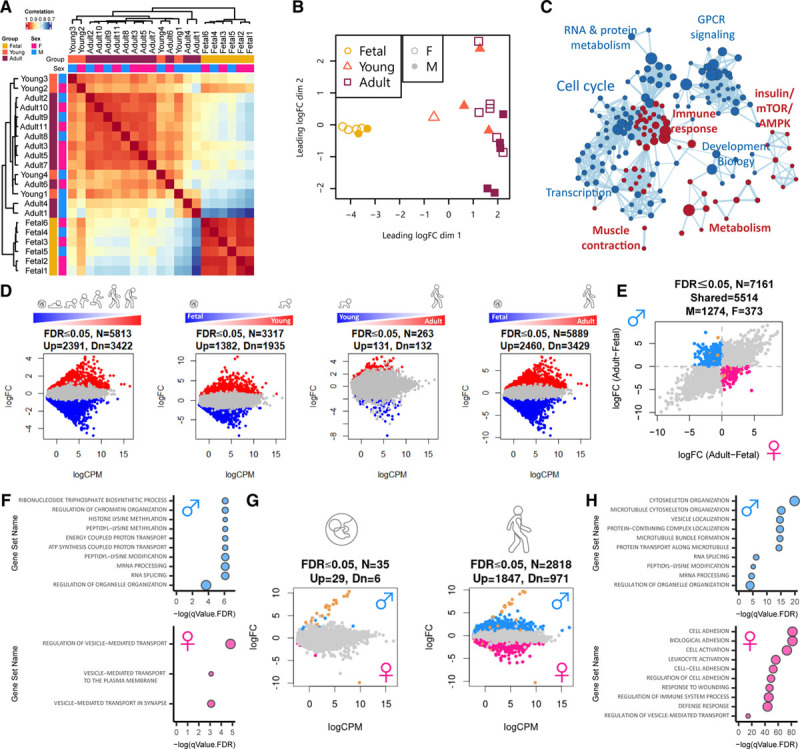
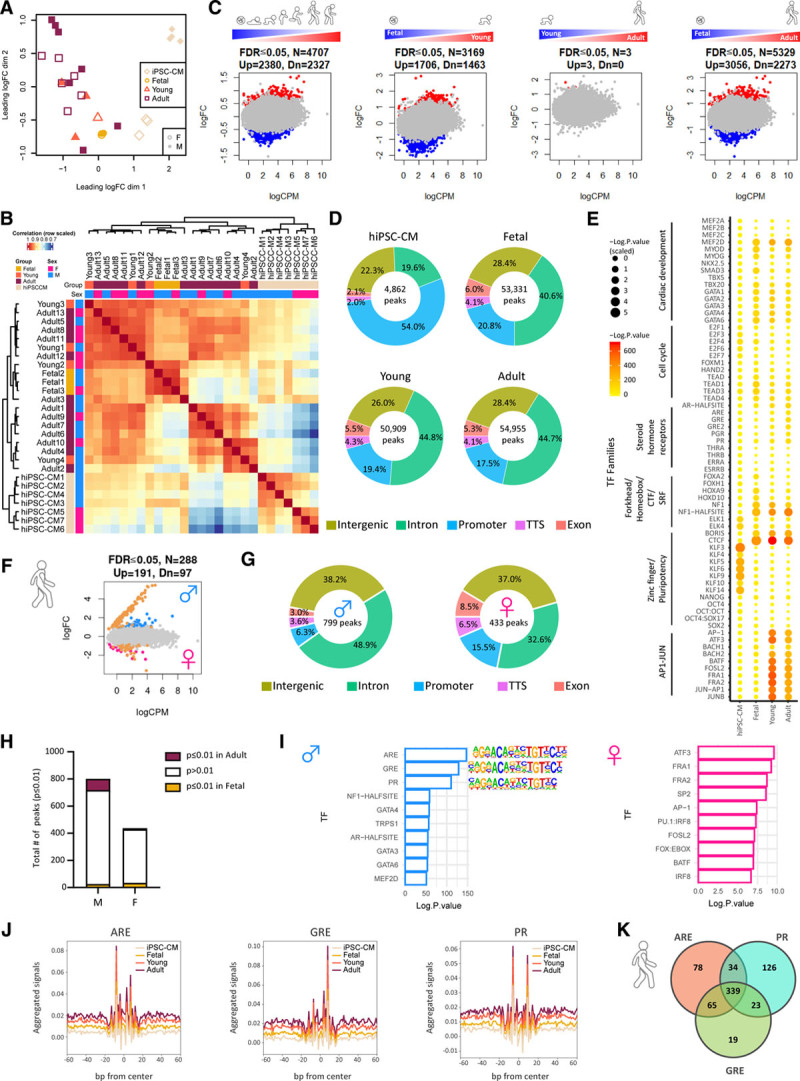
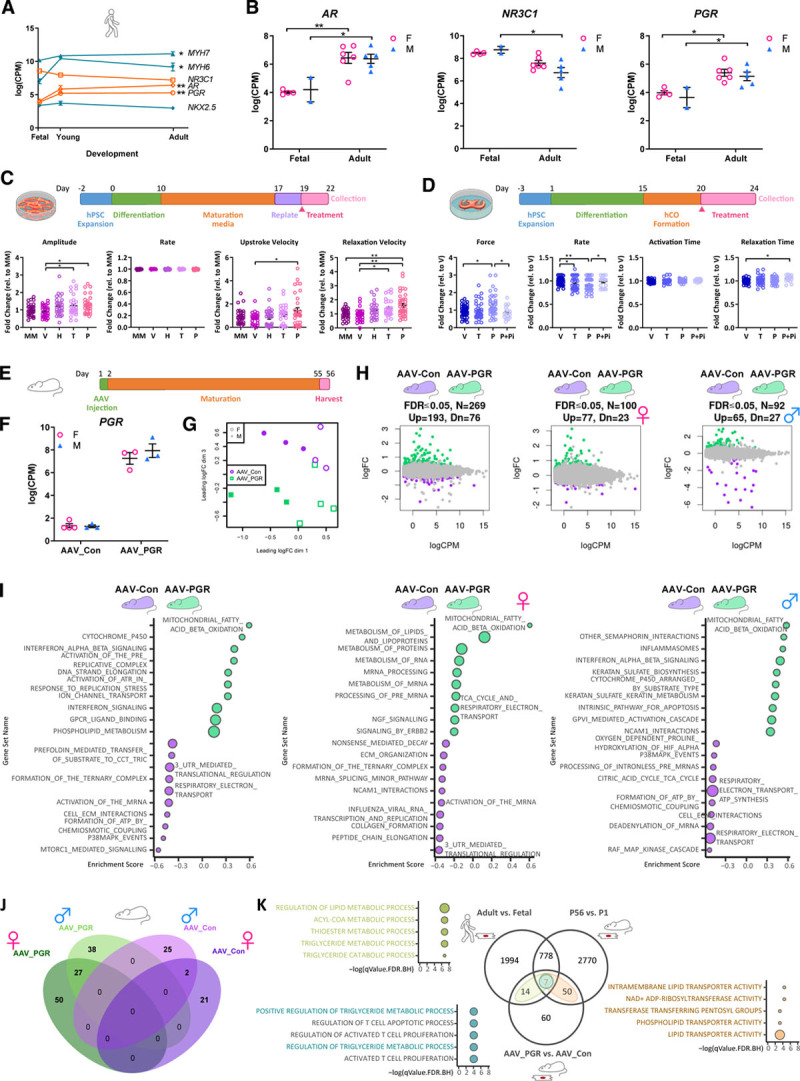
Methods: Single-nucleus RNA sequencing of 54 140 nuclei from 9 human donors was used to profile transcriptional changes in diverse cardiac cell types during maturation from fetal stages to adulthood. Bulk RNA sequencing and the Assay for Transposase-Accessible Chromatin using sequencing were used to further validate transcriptional changes and to profile alterations in the chromatin accessibility landscape in purified cardiomyocyte nuclei from 21 human donors. Functional validation studies of sex steroids implicated in cardiac maturation were performed in human pluripotent stem cell-derived cardiac organoids and mice.
Results: Our data identify the progesterone receptor as a key mediator of sex-dependent transcriptional programs during cardiomyocyte maturation. Functional validation studies in human cardiac organoids and mice demonstrate that the progesterone receptor drives sex-specific metabolic programs and maturation of cardiac contractile properties.
Conclusions: These data provide a blueprint for understanding human heart maturation in both sexes and reveal an important role for the progesterone receptor in human heart development.
Keywords: chromatin; hormone; human development; progesterone; sexual maturation; transcription factor.
Figures
Comment in
-
Progesterone receptor controls heart maturation.Nat Rev Cardiol. 2021 Jun;18(6):387. doi: 10.1038/s41569-021-00546-z. Nat Rev Cardiol. 2021. PMID: 33742184 No abstract available.
References
-
- Cui Y, Zheng Y, Liu X, Yan L, Fan X, Yong J, Hu Y, Dong J, Li Q, Wu X, et al. Single-cell transcriptome analysis maps the developmental track of the human heart. Cell Rep. 2019;26:1934–1950.e5. doi: 10.1016/j.celrep.2019.01.079 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
