

Cholesterol alters mitophagy by impairing optineurin recruitment and lysosomal clearance in Alzheimer's disease
- PMID: 33685483
- PMCID: PMC7941983
- DOI: 10.1186/s13024-021-00435-6
Cholesterol alters mitophagy by impairing optineurin recruitment and lysosomal clearance in Alzheimer's disease
Abstract
Background: Emerging evidence indicates that impaired mitophagy-mediated clearance of defective mitochondria is a critical event in Alzheimer's disease (AD) pathogenesis. Amyloid-beta (Aβ) metabolism and the microtubule-associated protein tau have been reported to regulate key components of the mitophagy machinery. However, the mechanisms that lead to mitophagy dysfunction in AD are not fully deciphered. We have previously shown that intraneuronal cholesterol accumulation can disrupt the autophagy flux, resulting in low Aβ clearance. In this study, we examine the impact of neuronal cholesterol changes on mitochondrial removal by autophagy.
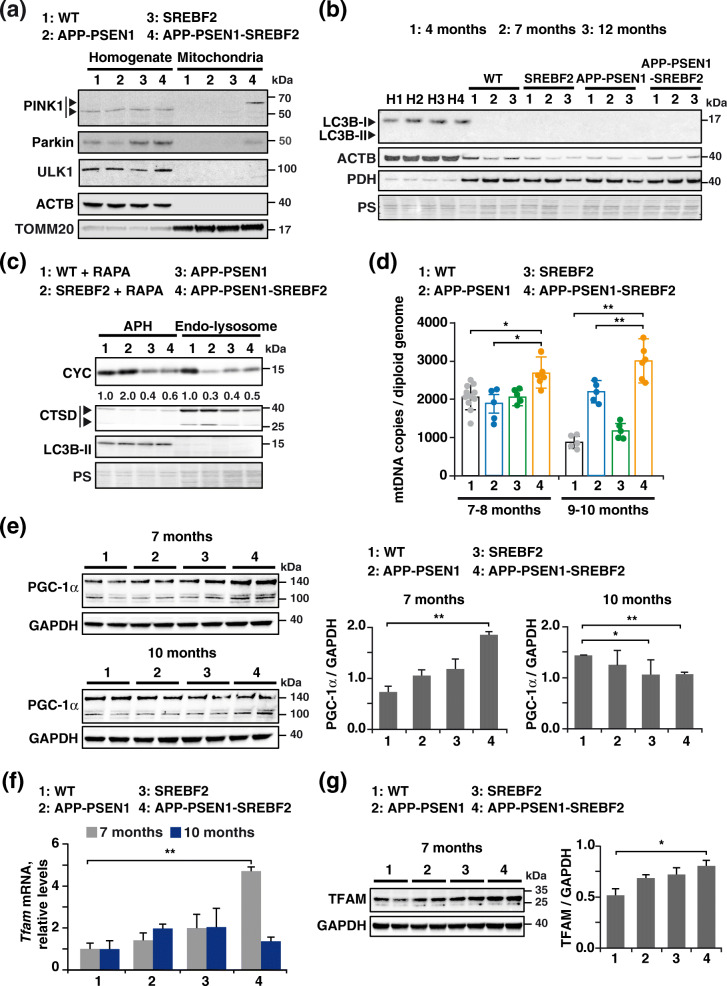
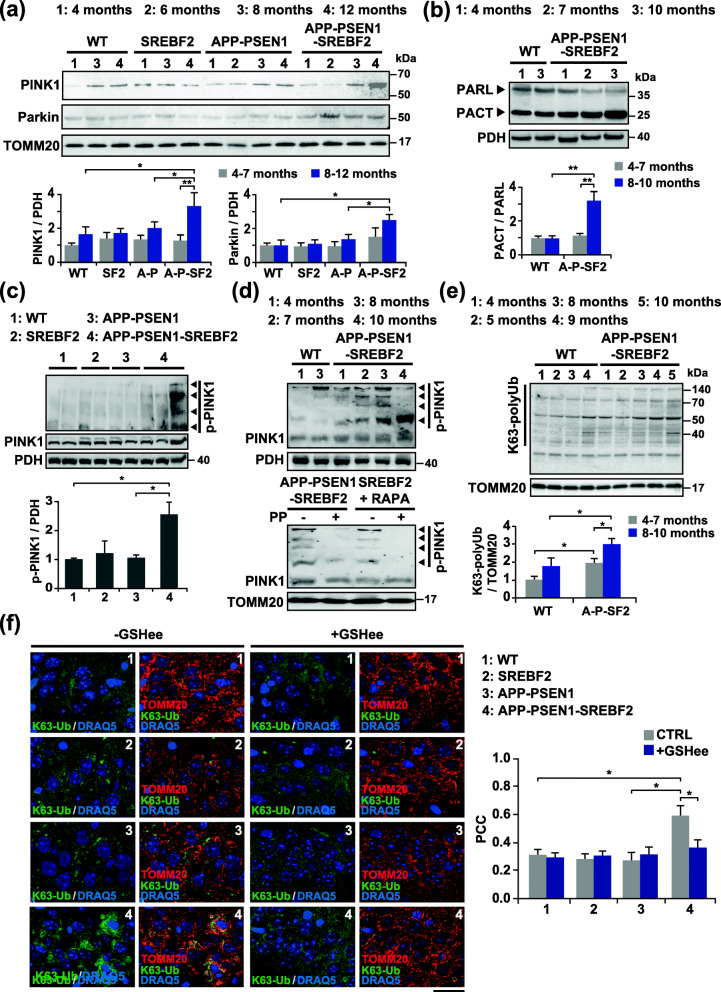
Methods: Regulation of PINK1-parkin-mediated mitophagy was investigated in conditions of acute (in vitro) and chronic (in vivo) high cholesterol loading using cholesterol-enriched SH-SY5Y cells, cultured primary neurons from transgenic mice overexpressing active SREBF2 (sterol regulatory element binding factor 2), and mice of increasing age that express the amyloid precursor protein with the familial Alzheimer Swedish mutation (Mo/HuAPP695swe) and mutant presenilin 1 (PS1-dE9) together with active SREBF2.
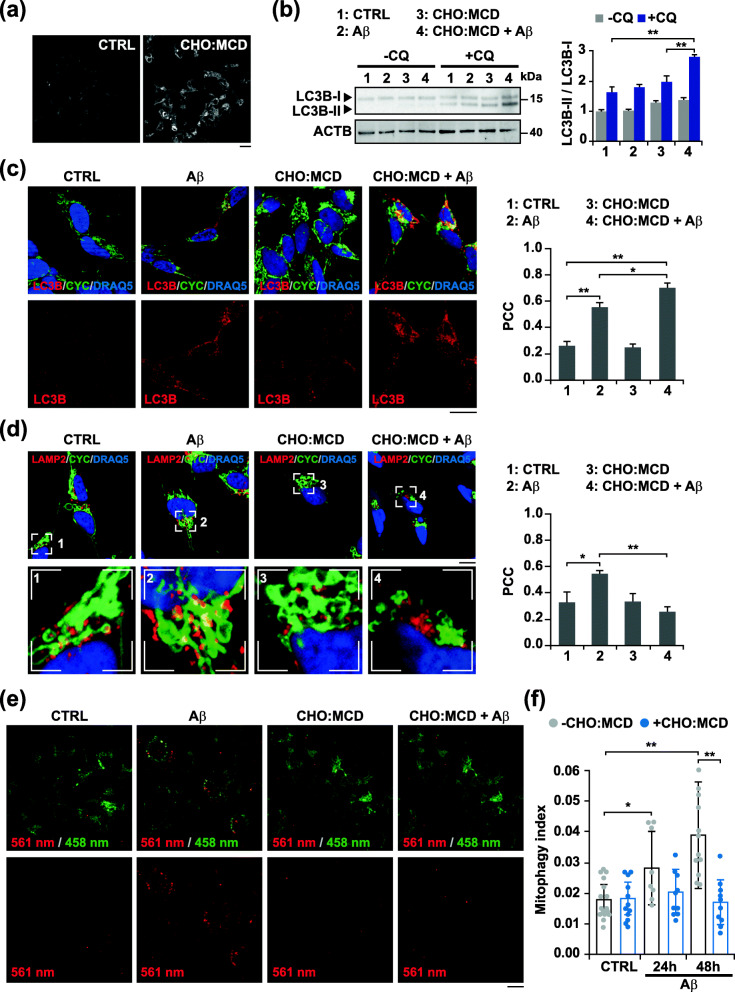
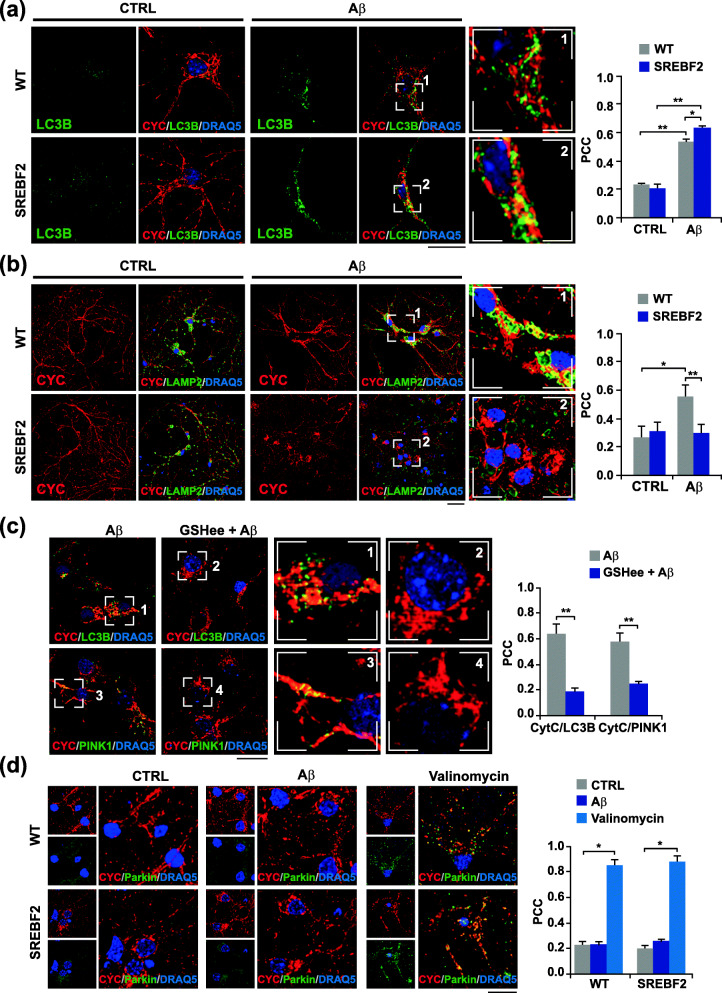
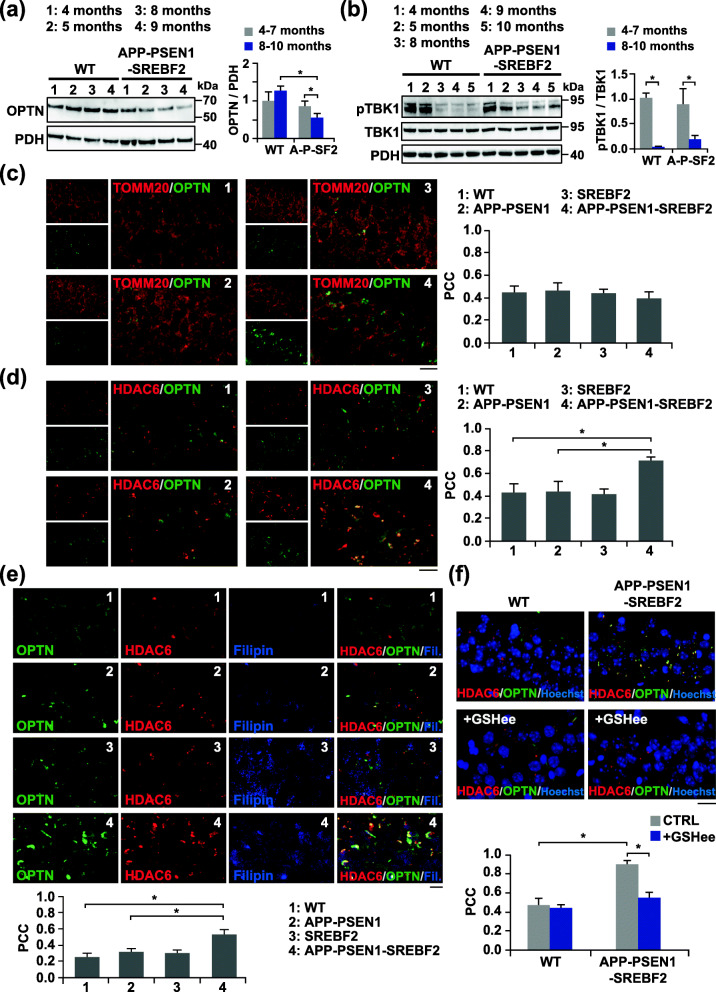
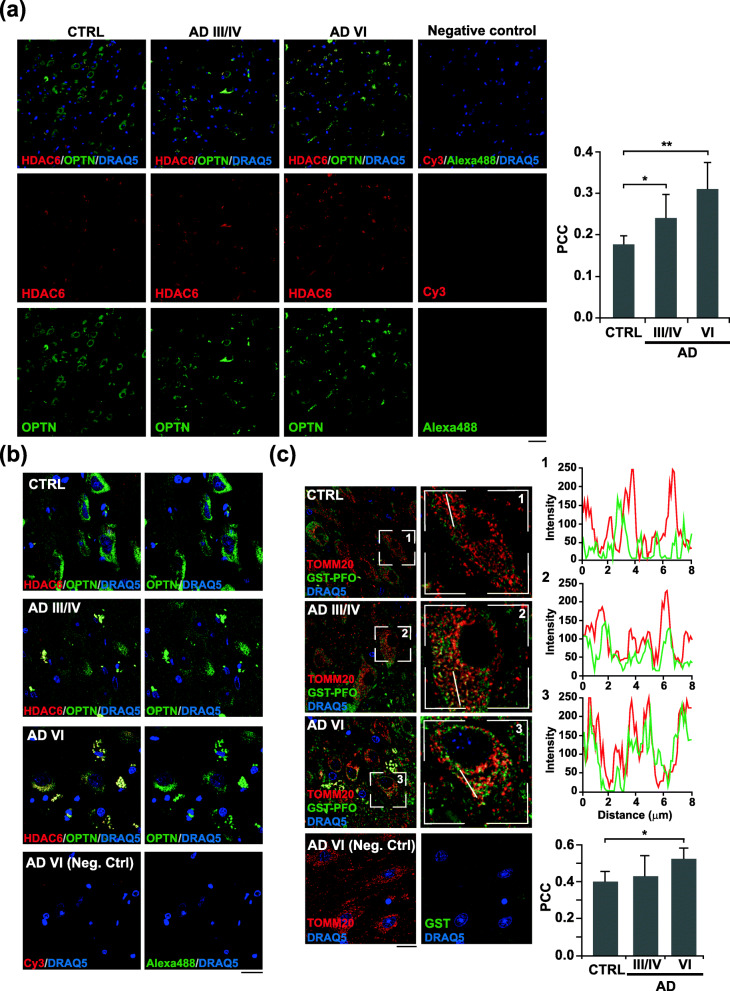
Results: In cholesterol-enriched SH-SY5Y cells and cultured primary neurons, high intracellular cholesterol levels stimulated mitochondrial PINK1 accumulation and mitophagosomes formation triggered by Aβ while impairing lysosomal-mediated clearance. Antioxidant recovery of cholesterol-induced mitochondrial glutathione (GSH) depletion prevented mitophagosomes formation indicating mitochondrial ROS involvement. Interestingly, when brain cholesterol accumulated chronically in aged APP-PSEN1-SREBF2 mice the mitophagy flux was affected at the early steps of the pathway, with defective recruitment of the key autophagy receptor optineurin (OPTN). Sustained cholesterol-induced alterations in APP-PSEN1-SREBF2 mice promoted an age-dependent accumulation of OPTN into HDAC6-positive aggresomes, which disappeared after in vivo treatment with GSH ethyl ester (GSHee). The analyses in post-mortem brain tissues from individuals with AD confirmed these findings, showing OPTN in aggresome-like structures that correlated with high mitochondrial cholesterol levels in late AD stages.
Conclusions: Our data demonstrate that accumulation of intracellular cholesterol reduces the clearance of defective mitochondria and suggest recovery of the cholesterol homeostasis and the mitochondrial scavenging of ROS as potential therapeutic targets for AD.
Keywords: APP-PSEN1 mice; Aggressomes; Glutathione; Mitochondria; Optineurin; Oxidative stress; PINK1; Parkin.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
