

Molecular Evaluation of Endoplasmic Reticulum Homeostasis Meets Humoral Immunity
- PMID: 33685797
- PMCID: PMC7611675
- DOI: 10.1016/j.tcb.2021.02.004
Molecular Evaluation of Endoplasmic Reticulum Homeostasis Meets Humoral Immunity
Abstract
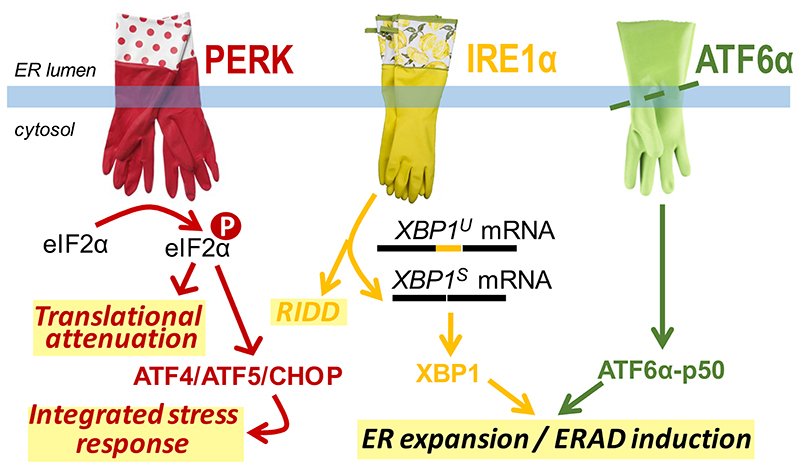
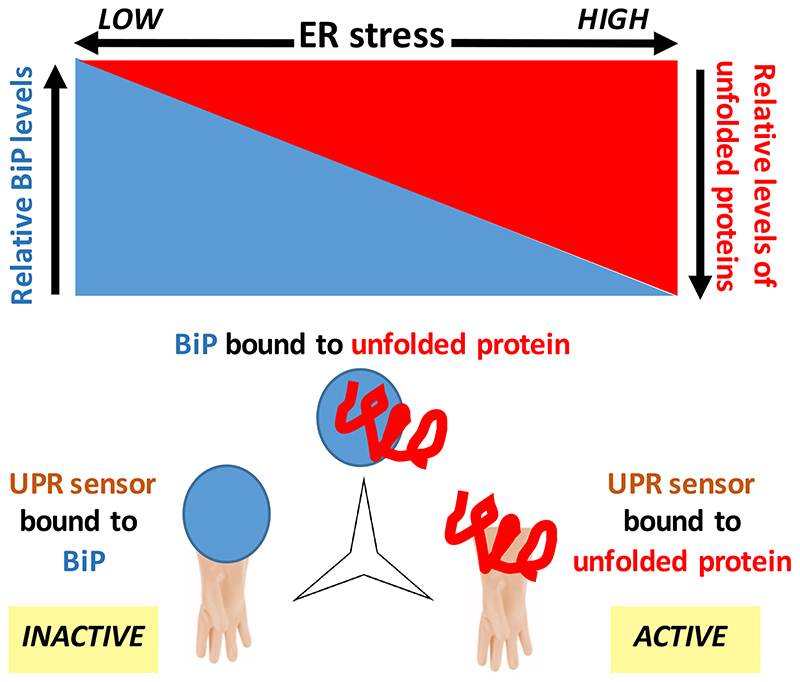
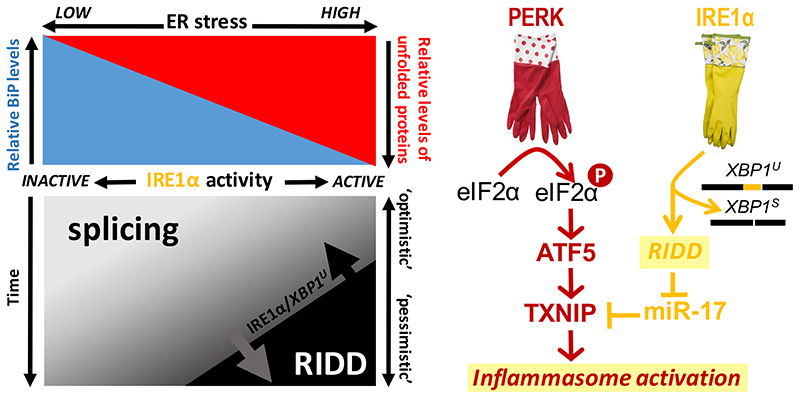
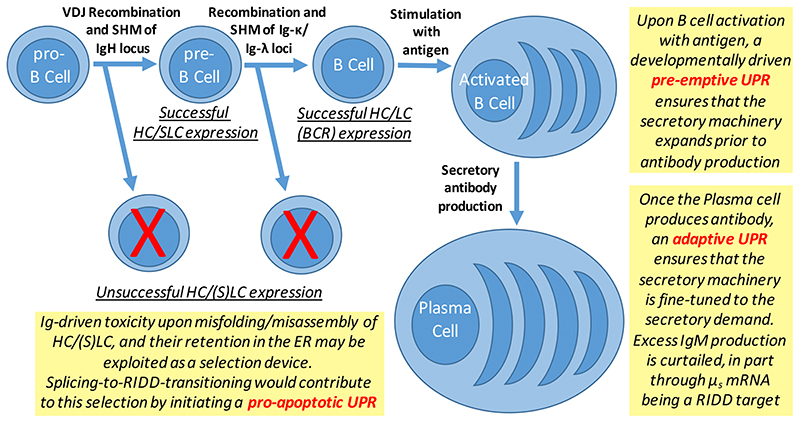
The biosynthesis of about one third of the human proteome, including membrane receptors and secreted proteins, occurs in the endoplasmic reticulum (ER). Conditions that perturb ER homeostasis activate the unfolded protein response (UPR). An 'optimistic' UPR output aims at restoring homeostasis by reinforcement of machineries that guarantee efficiency and fidelity of protein biogenesis in the ER. Yet, once the UPR 'deems' that ER homeostatic readjustment fails, it transitions to a 'pessimistic' output, which, depending on the cell type, will result in apoptosis. In this article, we discuss emerging concepts on how the UPR 'evaluates' ER stress, how the UPR is repurposed, in particular in B cells, and how UPR-driven counter-selection of cells undergoing homeostatic failure serves organismal homeostasis and humoral immunity.
Keywords: B cell development; RIDD; antibody production; endoplasmic reticulum; proteostasis; unfolded protein response.
Copyright © 2021 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
