

Proteomic and transcriptomic profiling reveal different aspects of aging in the kidney
- PMID: 33687326
- PMCID: PMC8096428
- DOI: 10.7554/eLife.62585
Proteomic and transcriptomic profiling reveal different aspects of aging in the kidney
Abstract
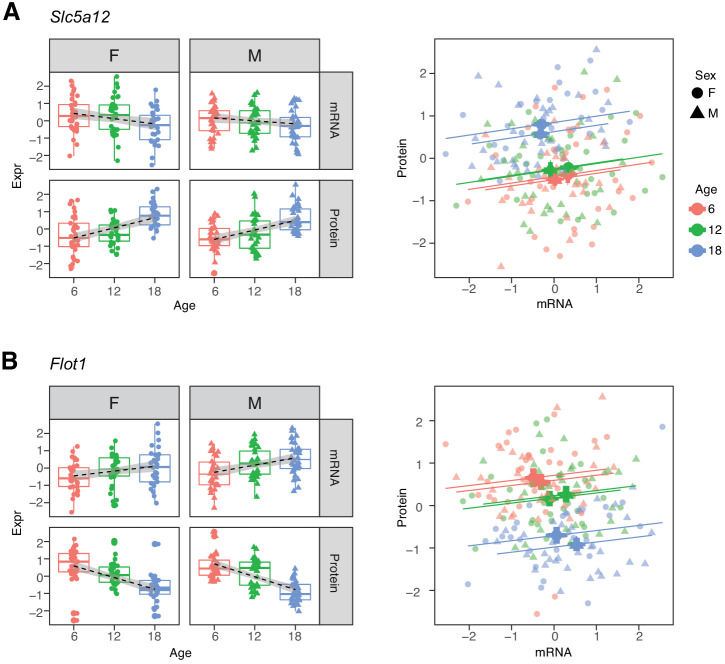
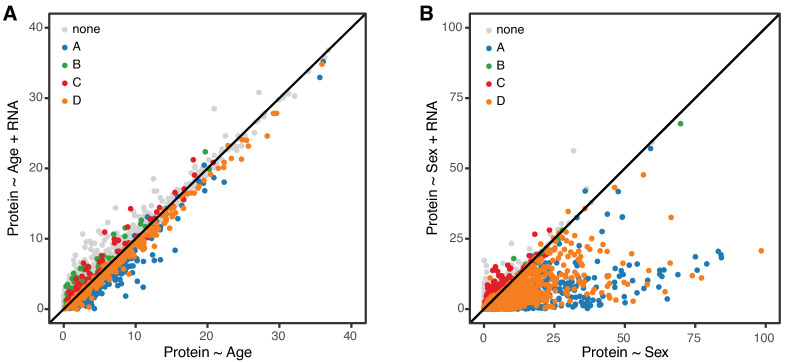
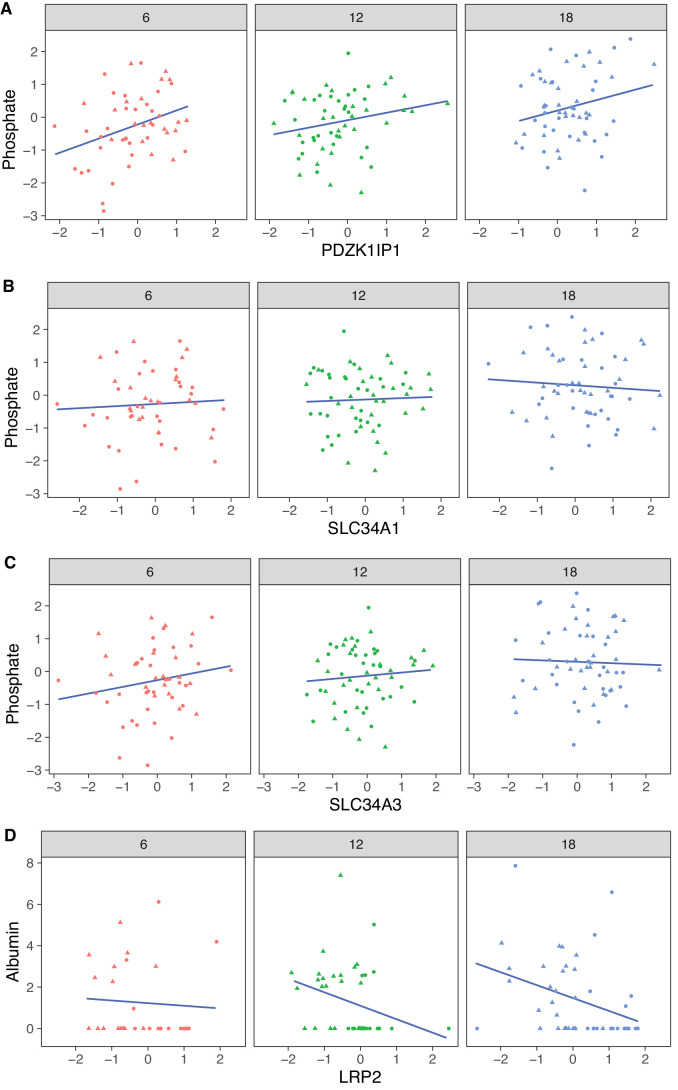
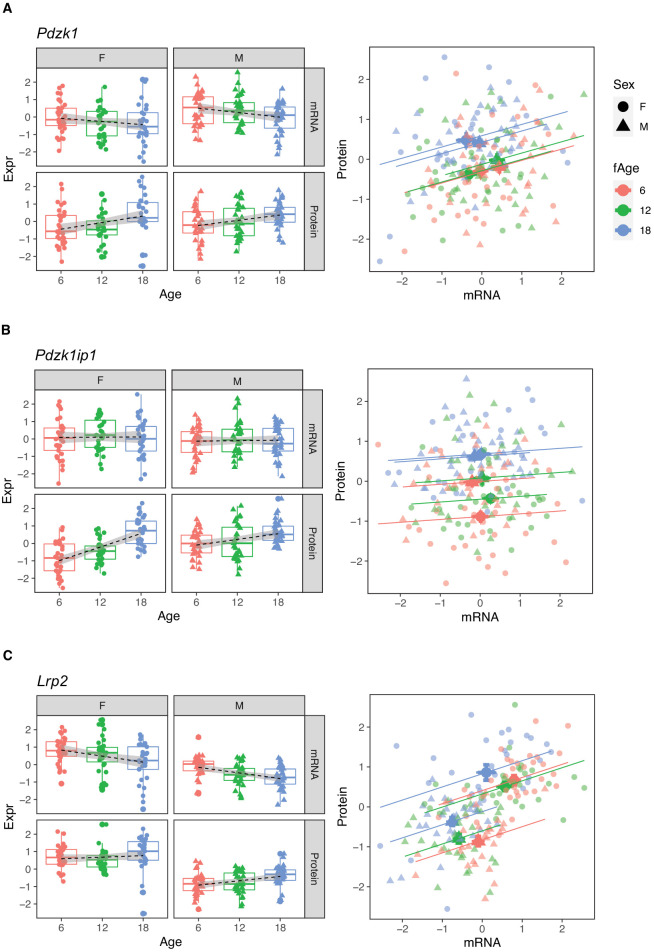
Little is known about the molecular changes that take place in the kidney during the aging process. In order to better understand these changes, we measured mRNA and protein levels in genetically diverse mice at different ages. We observed distinctive change in mRNA and protein levels as a function of age. Changes in both mRNA and protein are associated with increased immune infiltration and decreases in mitochondrial function. Proteins show a greater extent of change and reveal changes in a wide array of biological processes including unique, organ-specific features of aging in kidney. Most importantly, we observed functionally important age-related changes in protein that occur in the absence of corresponding changes in mRNA. Our findings suggest that mRNA profiling alone provides an incomplete picture of molecular aging in the kidney and that examination of changes in proteins is essential to understand aging processes that are not transcriptionally regulated.
Keywords: aging; computational biology; kidney; mRNA; mouse; protein; systems biology.
© 2021, Takemon et al.
Conflict of interest statement
YT, JC, IG, DS, OD, SG, GC, RK No competing interests declared
Figures
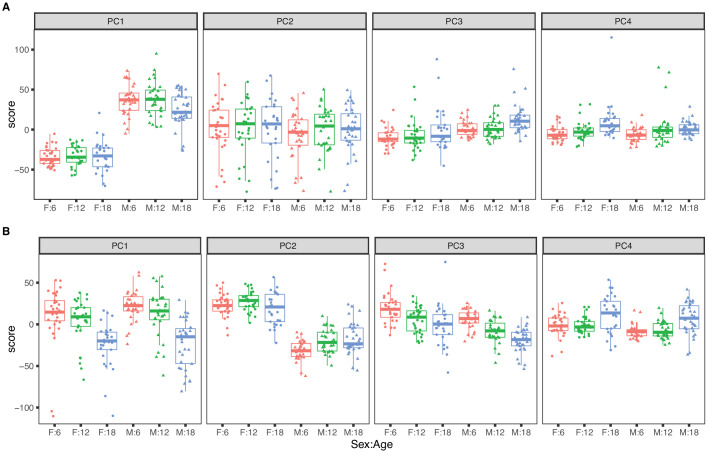
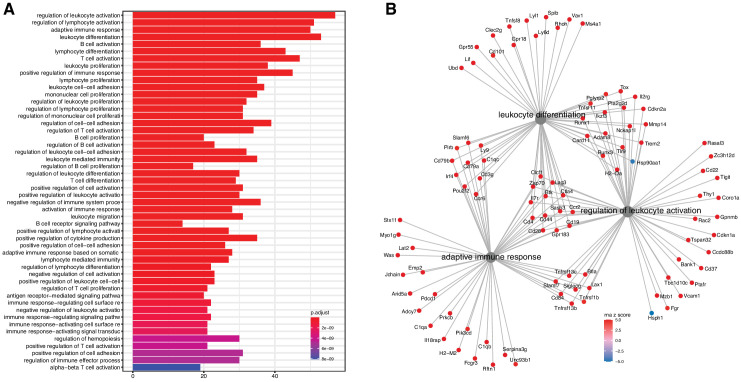
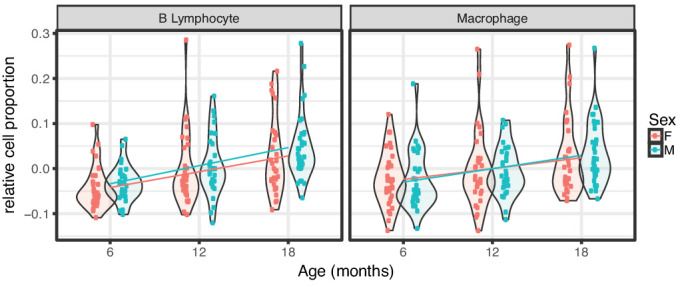
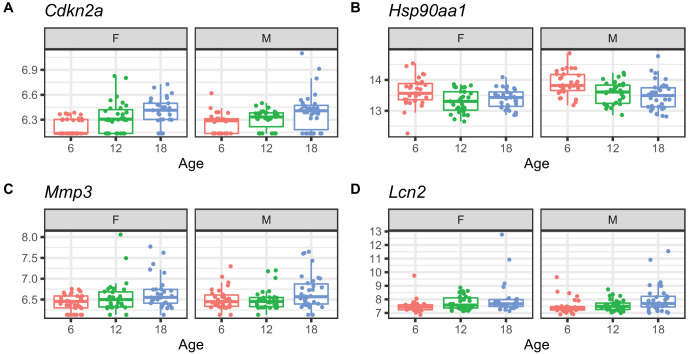
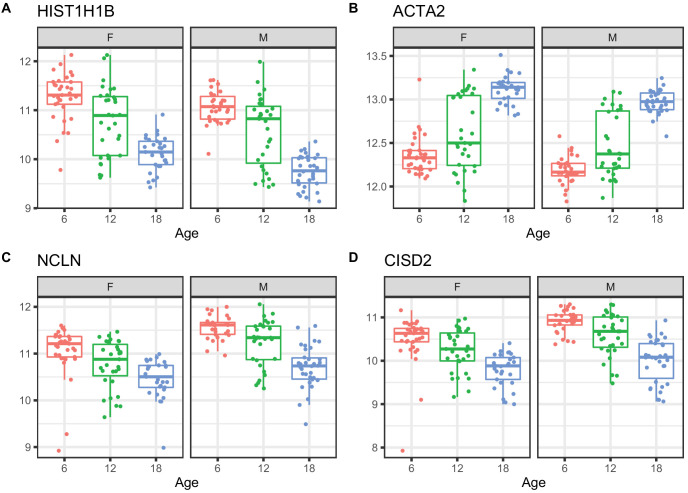
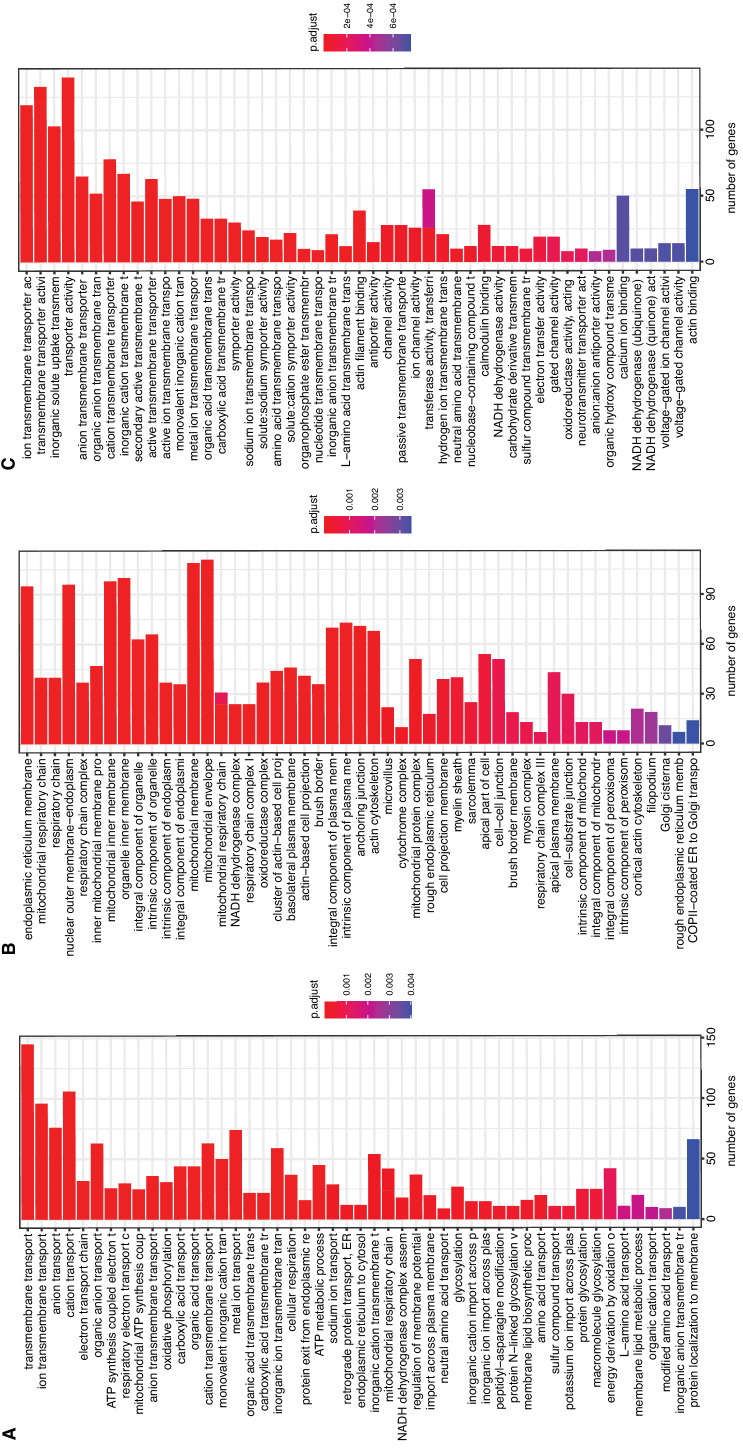
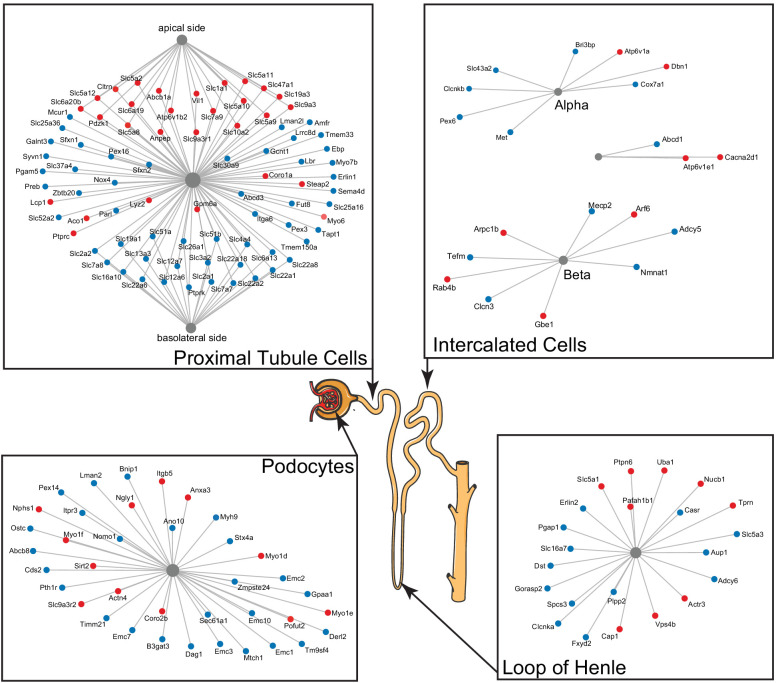
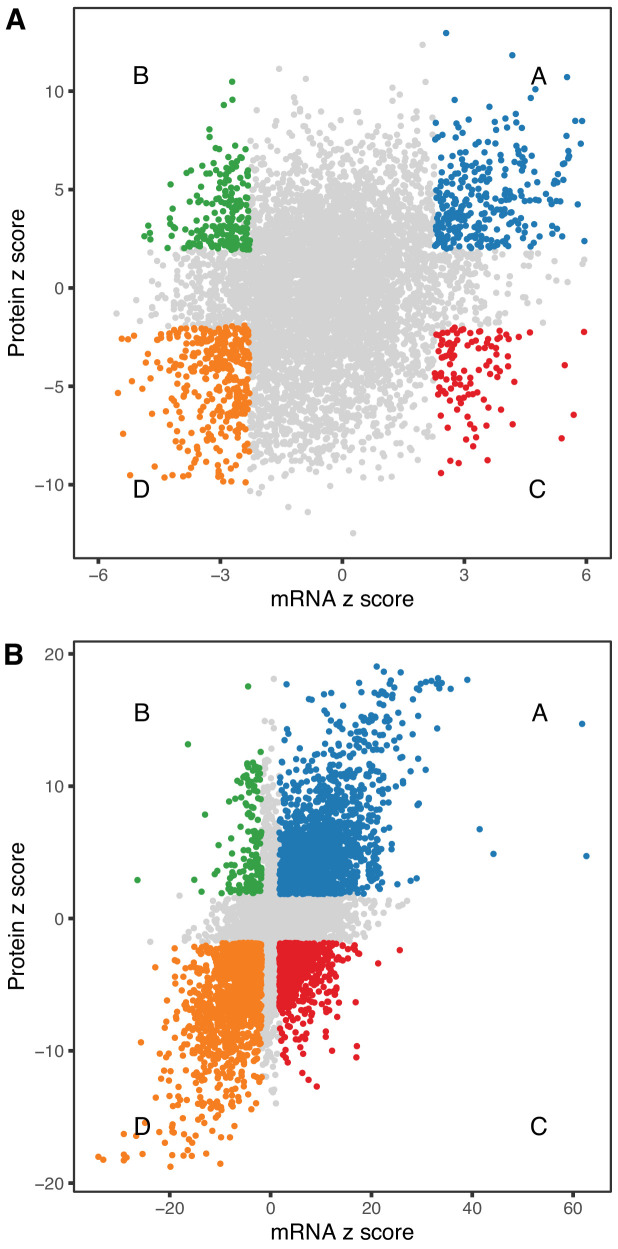
References
-
- Almanzar N, Antony J, Baghel A, Bakerman I, Bansal I, Barres B, Beachy P, Berdnik D, Bilen B, Brownfield D, Cain C, Chan C, Chen M, Clarke M, Conley S, Darmanis S, Demers A, Demir K, Morree A, Divita T, Bois H, Ebadi H, Espinoza F, Fish M. A single-cell transcriptomic atlas characterizes ageing tissues in the mouse. GanNature. 2020;583:590–595. doi: 10.1038/s41586-020-2496-1. - DOI - PMC - PubMed
-
- Bolignano D, Lacquaniti A, Coppolino G, Donato V, Campo S, Fazio MR, Nicocia G, Buemi M. Neutrophil Gelatinase-Associated lipocalin (NGAL) and progression of chronic kidney disease. Clinical Journal of the American Society of Nephrology. 2009;4:337–344. doi: 10.2215/CJN.03530708. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
