

Genomic analyses provide insights into peach local adaptation and responses to climate change
- PMID: 33687945
- PMCID: PMC8015852
- DOI: 10.1101/gr.261032.120
Genomic analyses provide insights into peach local adaptation and responses to climate change
Abstract
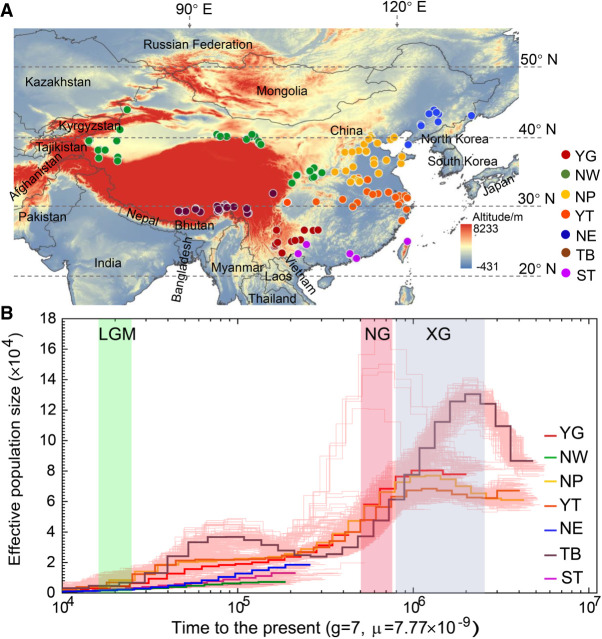
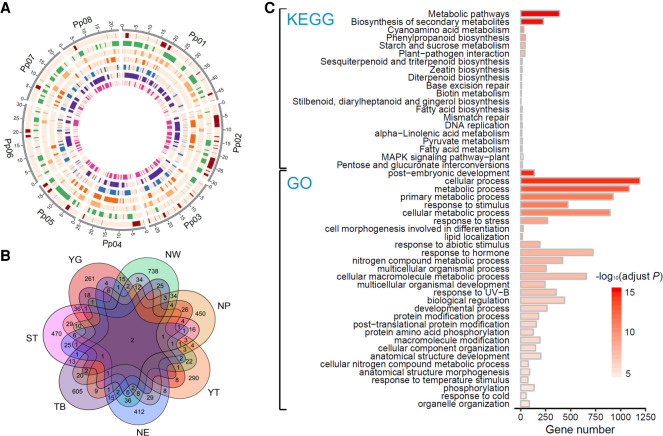
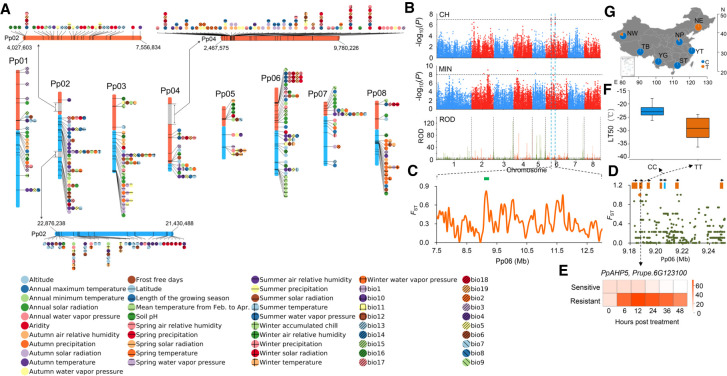
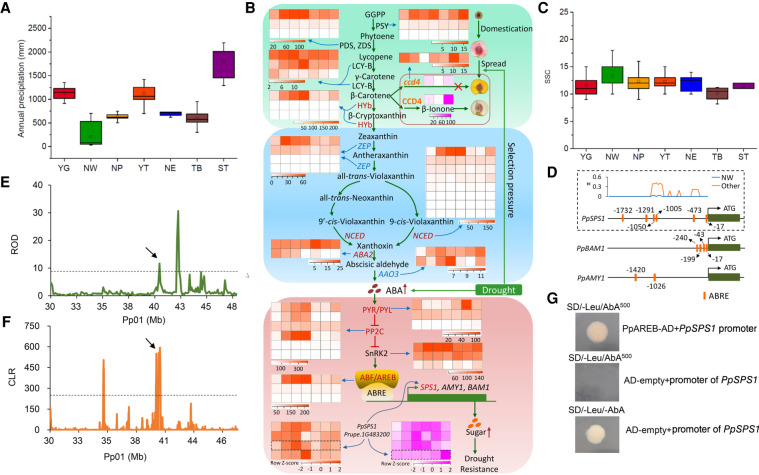
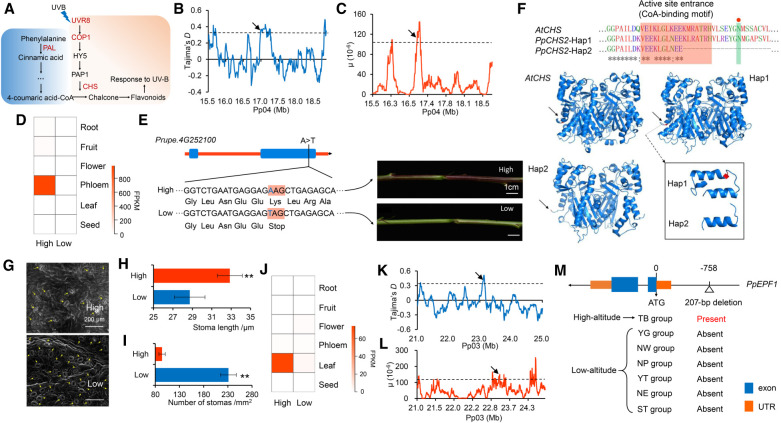
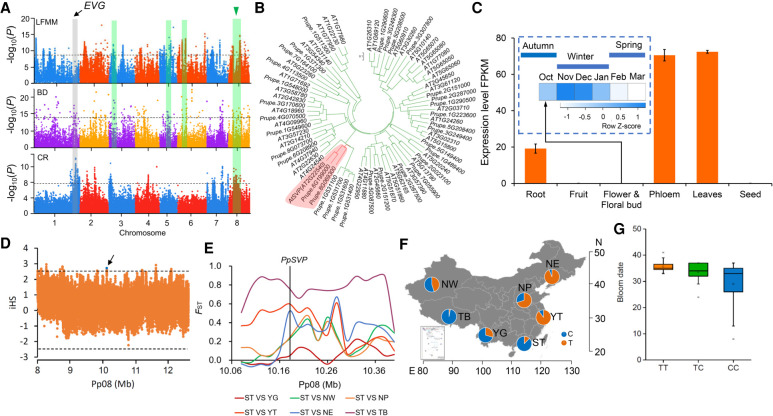
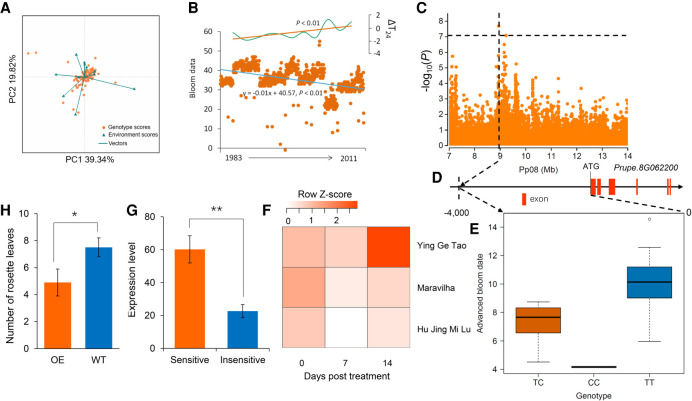
The environment has constantly shaped plant genomes, but the genetic bases underlying how plants adapt to environmental influences remain largely unknown. We constructed a high-density genomic variation map of 263 geographically representative peach landraces and wild relatives. A combination of whole-genome selection scans and genome-wide environmental association studies (GWEAS) was performed to reveal the genomic bases of peach adaptation to diverse climates. A total of 2092 selective sweeps that underlie local adaptation to both mild and extreme climates were identified, including 339 sweeps conferring genomic pattern of adaptation to high altitudes. Using genome-wide environmental association studies (GWEAS), a total of 2755 genomic loci strongly associated with 51 specific environmental variables were detected. The molecular mechanism underlying adaptive evolution of high drought, strong UVB, cold hardiness, sugar content, flesh color, and bloom date were revealed. Finally, based on 30 yr of observation, a candidate gene associated with bloom date advance, representing peach responses to global warming, was identified. Collectively, our study provides insights into molecular bases of how environments have shaped peach genomes by natural selection and adds candidate genes for future studies on evolutionary genetics, adaptation to climate changes, and breeding.
© 2021 Li et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
-
- Annicchiarico P. 1997. Additive main effects and multiplicative interaction (AMMI) analysis of genotype-location interaction in variety trials repeated over years. Theor Appl Genet 94: 1072–1077. 10.1007/s001220050517 - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical