

cMyBPC phosphorylation modulates the effect of omecamtiv mecarbil on myocardial force generation
- PMID: 33688929
- PMCID: PMC7953254
- DOI: 10.1085/jgp.202012816
cMyBPC phosphorylation modulates the effect of omecamtiv mecarbil on myocardial force generation
Abstract
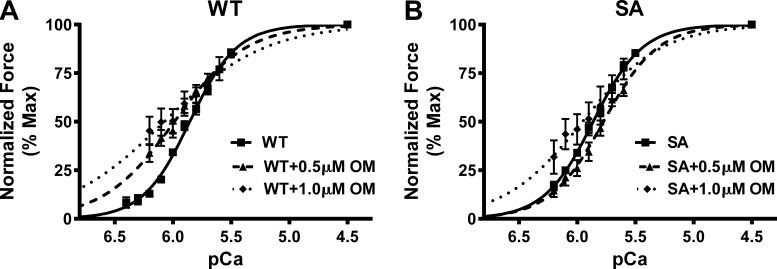
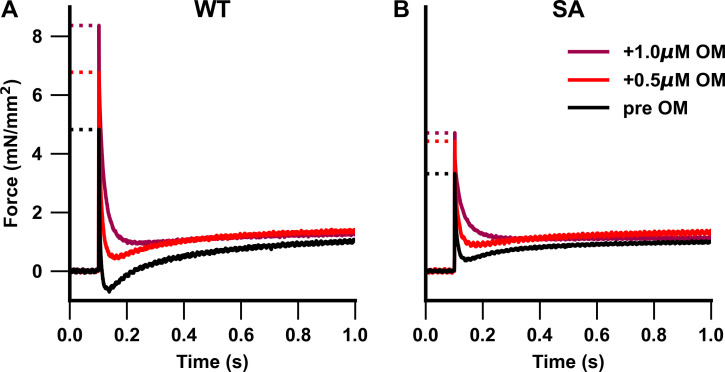
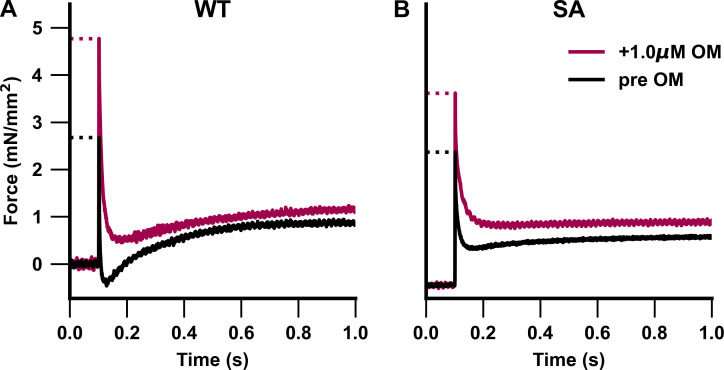
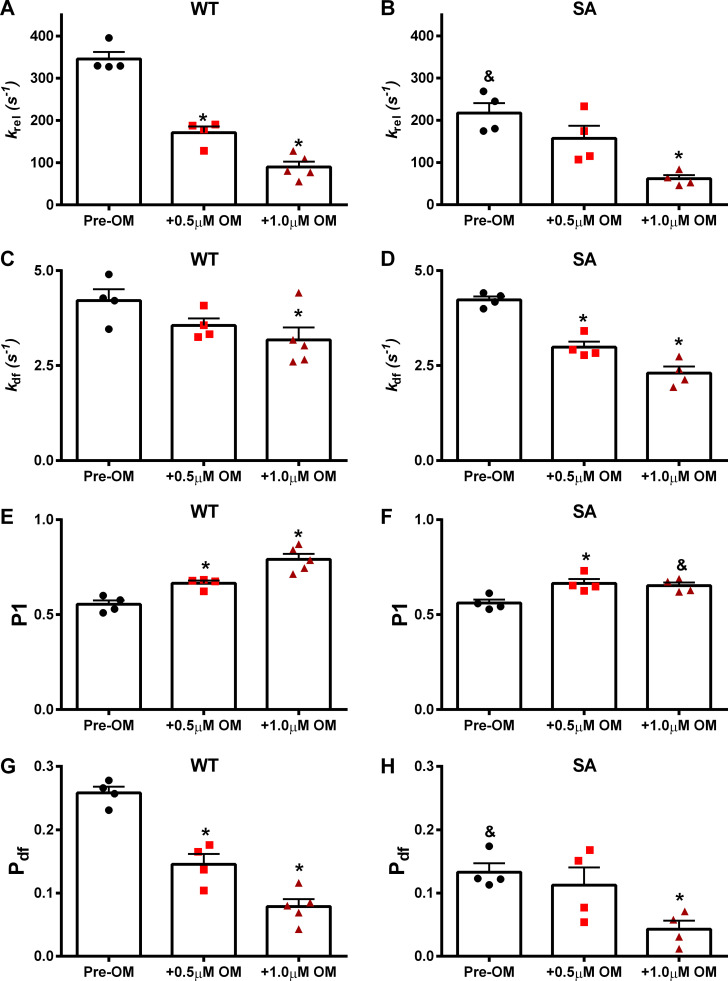
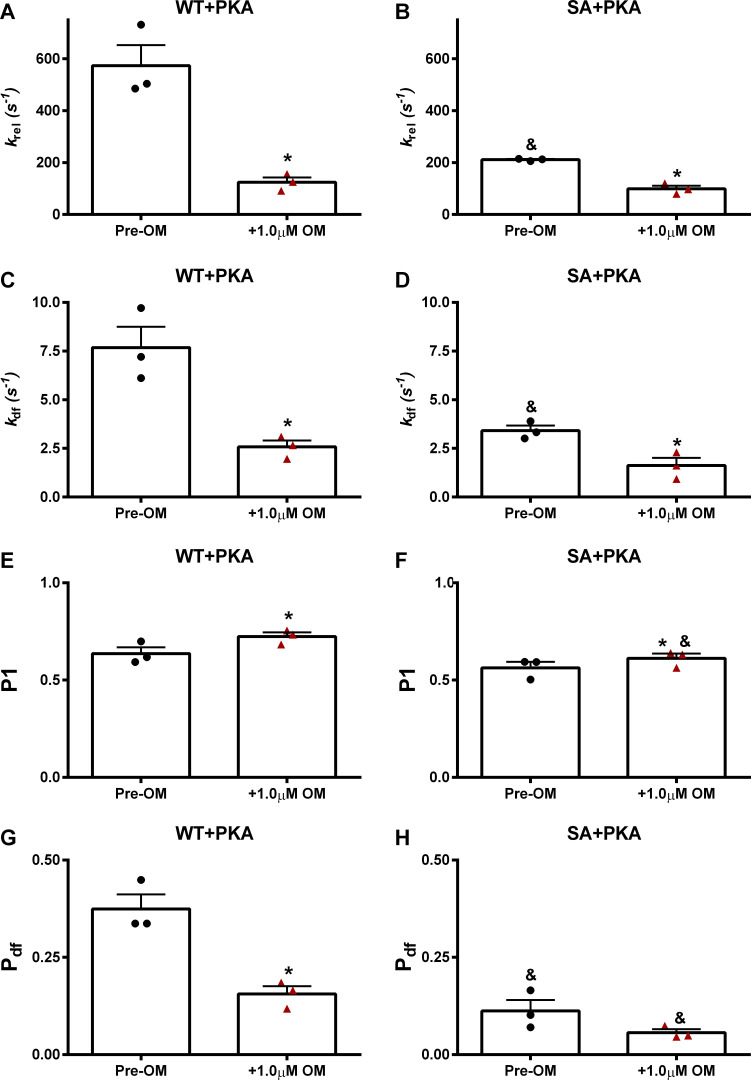
Omecamtiv mecarbil (OM), a direct myosin motor activator, is currently being tested as a therapeutic replacement for conventional inotropes in heart failure (HF) patients. It is known that HF patients exhibit dysregulated β-adrenergic signaling and decreased cardiac myosin-binding protein C (cMyBPC) phosphorylation, a critical modulator of myocardial force generation. However, the functional effects of OM in conditions of altered cMyBPC phosphorylation have not been established. Here, we tested the effects of OM on force generation and cross-bridge (XB) kinetics using murine myocardial preparations isolated from wild-type (WT) hearts and from hearts expressing S273A, S282A, and S302A substitutions (SA) in the M domain, between the C1 and C2 domains of cMyBPC, which cannot be phosphorylated. At submaximal Ca2+ activations, OM-mediated force enhancements were less pronounced in SA than in WT myocardial preparations. Additionally, SA myocardial preparations lacked the dose-dependent increases in force that were observed in WT myocardial preparations. Following OM incubation, the basal differences in the rate of XB detachment (krel) between WT and SA myocardial preparations were abolished, suggesting that OM differentially affects the XB behavior when cMyBPC phosphorylation is reduced. Similarly, in myocardial preparations pretreated with protein kinase A to phosphorylate cMyBPC, incubation with OM significantly slowed krel in both the WT and SA myocardial preparations. Collectively, our data suggest there is a strong interplay between the effects of OM and XB behavior, such that it effectively uncouples the sarcomere from cMyBPC phosphorylation levels. Our findings imply that OM may significantly alter the in vivo cardiac response to β-adrenergic stimulation.
© 2021 Mamidi et al.
Figures
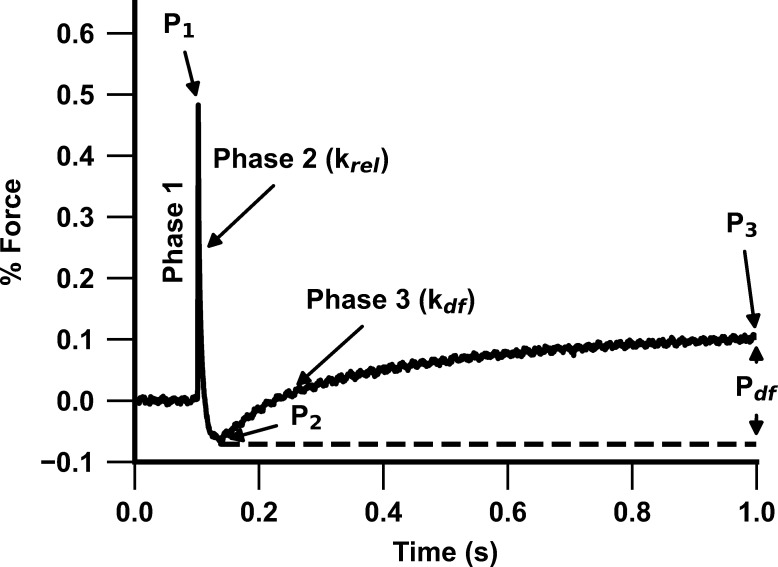
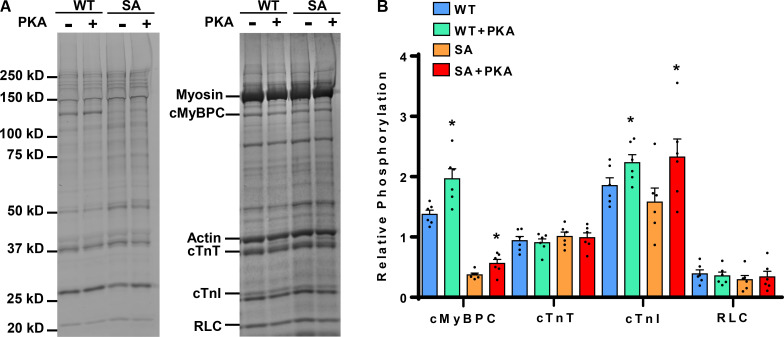
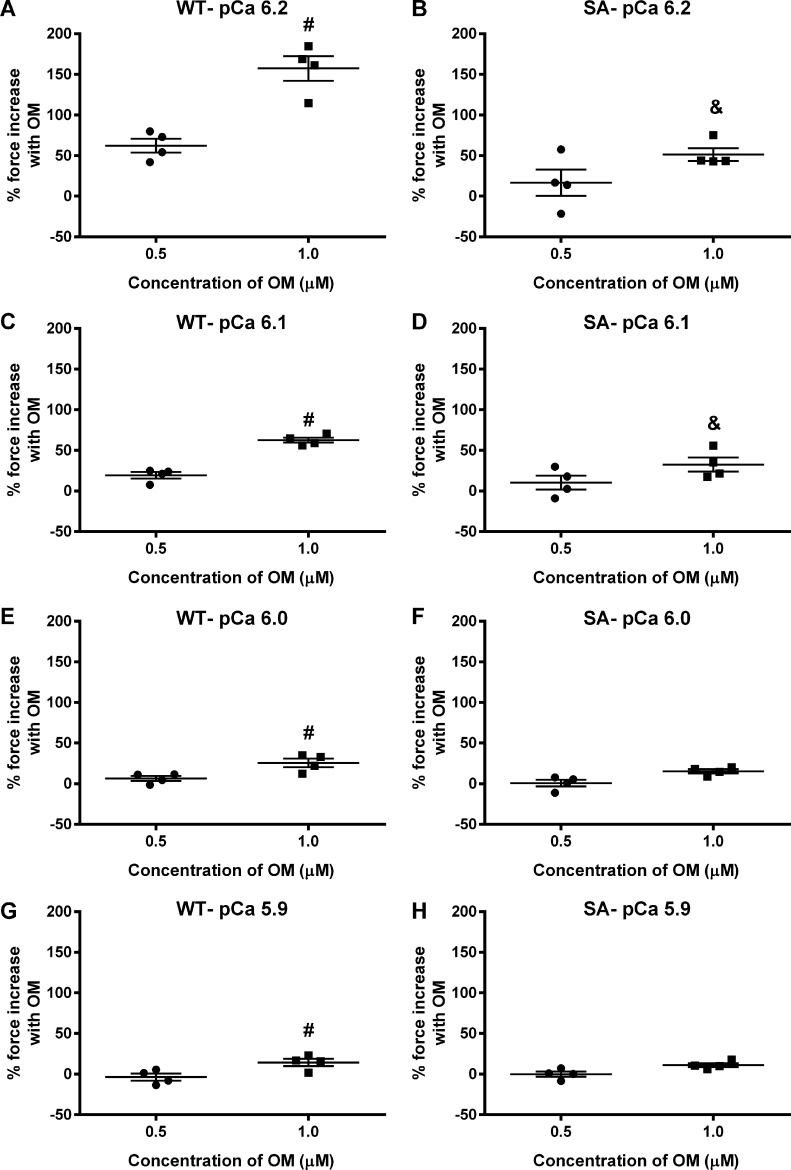
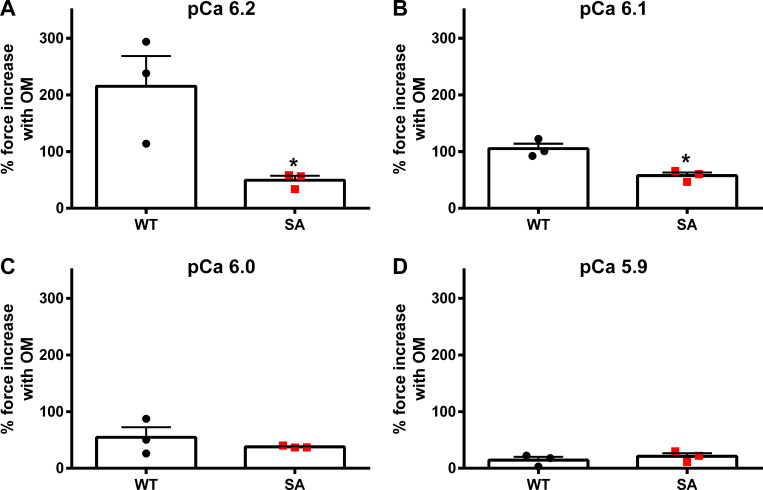
Comment in
-
cMyBPC phosphorylation alters response to heart failure drug.J Gen Physiol. 2021 Jul 5;153(7):e202112973. doi: 10.1085/jgp.202112973. Epub 2021 Jun 14. J Gen Physiol. 2021. PMID: 34125165 Free PMC article.
References
-
- Ahmad, T., Miller P.E., McCullough M., Desai N.R., Riello R., Psotka M., Böhm M., Allen L.A., Teerlink J.R., Rosano G.M.C., and Lindenfeld J.. 2019. Why has positive inotropy failed in chronic heart failure? Lessons from prior inotrope trials. Eur. J. Heart Fail. 21:1064–1078. 10.1002/ejhf.1557 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
