

Transcriptional bursts explain autosomal random monoallelic expression and affect allelic imbalance
- PMID: 33690599
- PMCID: PMC7978379
- DOI: 10.1371/journal.pcbi.1008772
Transcriptional bursts explain autosomal random monoallelic expression and affect allelic imbalance
Abstract
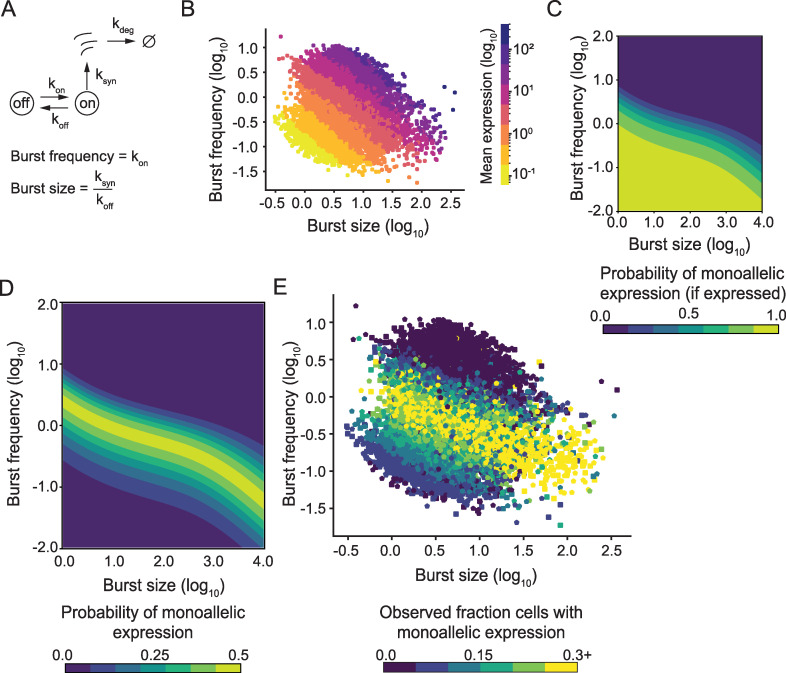
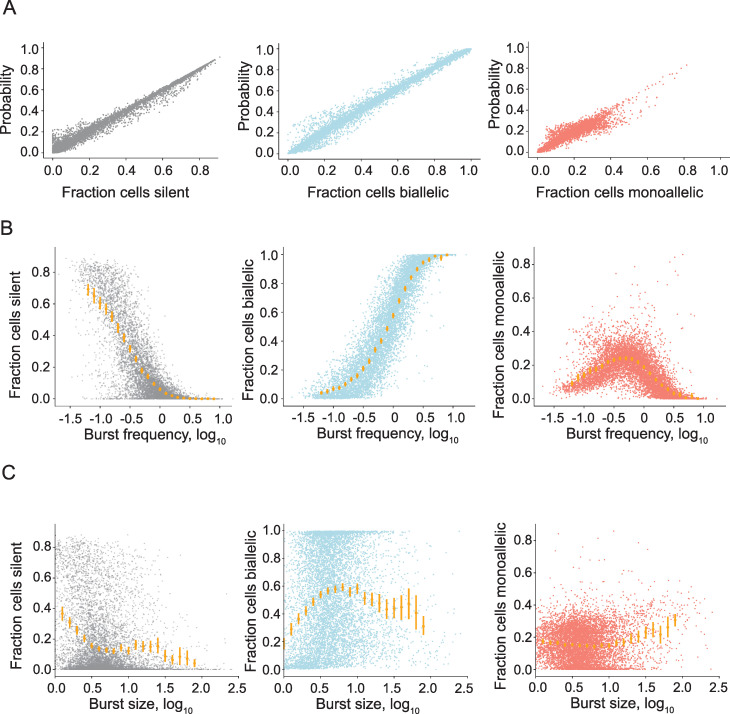
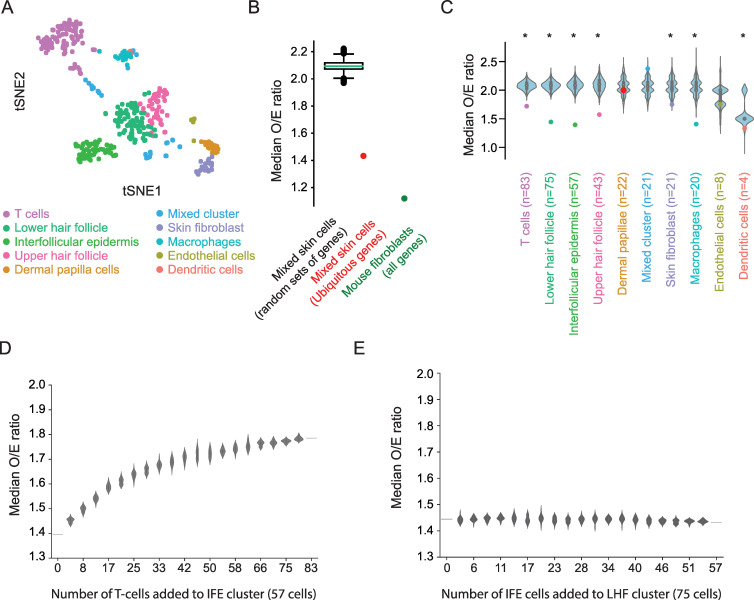
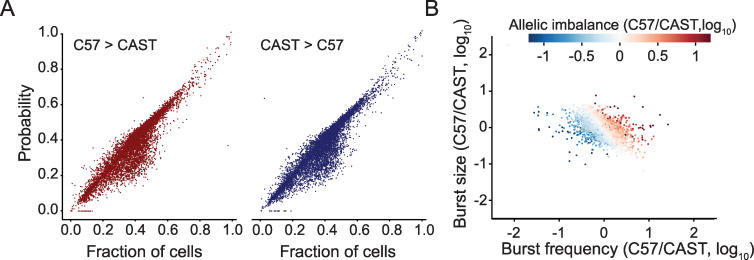
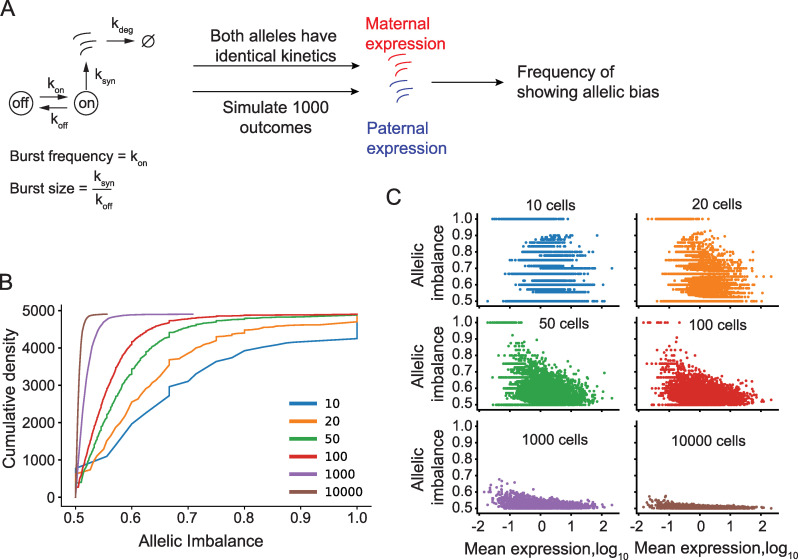
Transcriptional bursts render substantial biological noise in cellular transcriptomes. Here, we investigated the theoretical extent of allelic expression resulting from transcriptional bursting and how it compared to the amount biallelic, monoallelic and allele-biased expression observed in single-cell RNA-sequencing (scRNA-seq) data. We found that transcriptional bursting can explain the allelic expression patterns observed in single cells, including the frequent observations of autosomal monoallelic gene expression. Importantly, we identified that the burst frequency largely determined the fraction of cells with monoallelic expression, whereas the burst size had little effect on monoallelic observations. The high consistency between the bursting model predictions and scRNA-seq observations made it possible to assess the heterogeneity of a group of cells as their deviation in allelic observations from the expected. Finally, both burst frequency and size contributed to allelic imbalance observations and reinforced that studies of allelic imbalance can be confounded from the inherent noise in transcriptional bursting. Altogether, we demonstrate that allele-level transcriptional bursting renders widespread, although predictable, amounts of monoallelic and biallelic expression in single cells and cell populations.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
