

Blunted diurnal firing in lateral habenula projections to dorsal raphe nucleus and delayed photoentrainment in stress-susceptible mice
- PMID: 33690628
- PMCID: PMC7984642
- DOI: 10.1371/journal.pbio.3000709
Blunted diurnal firing in lateral habenula projections to dorsal raphe nucleus and delayed photoentrainment in stress-susceptible mice
Abstract
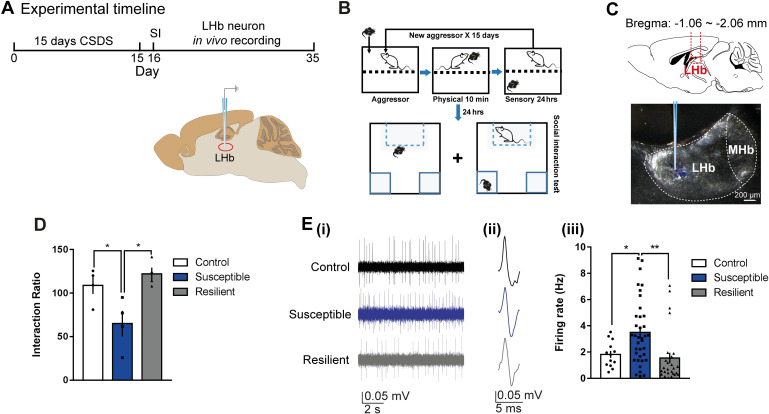
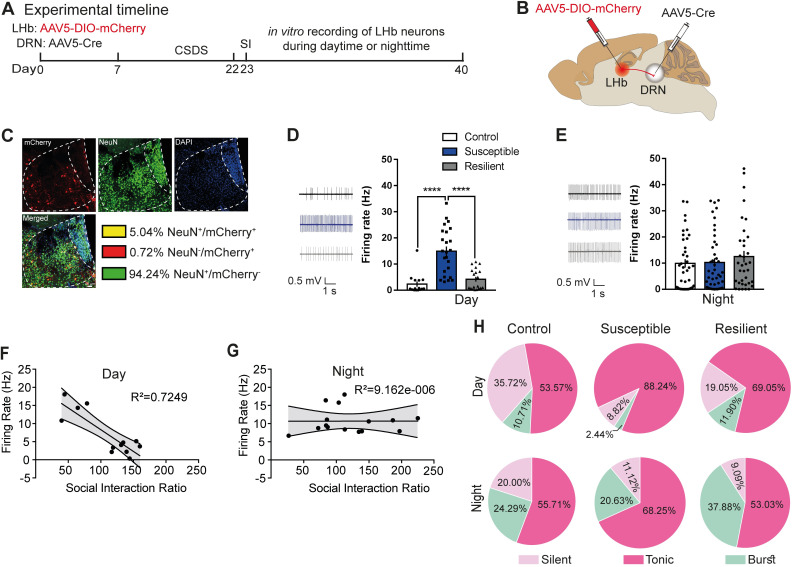
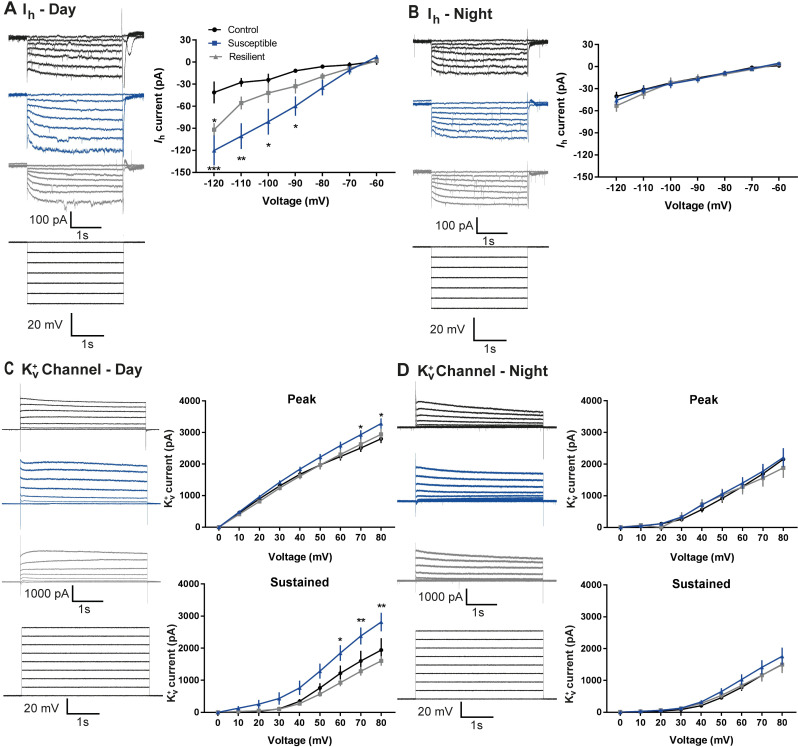
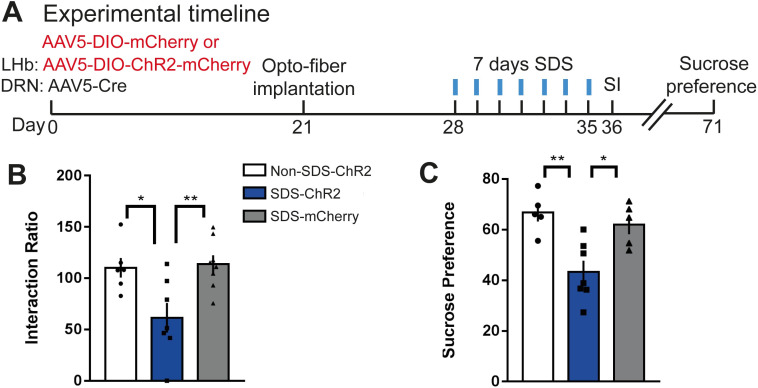
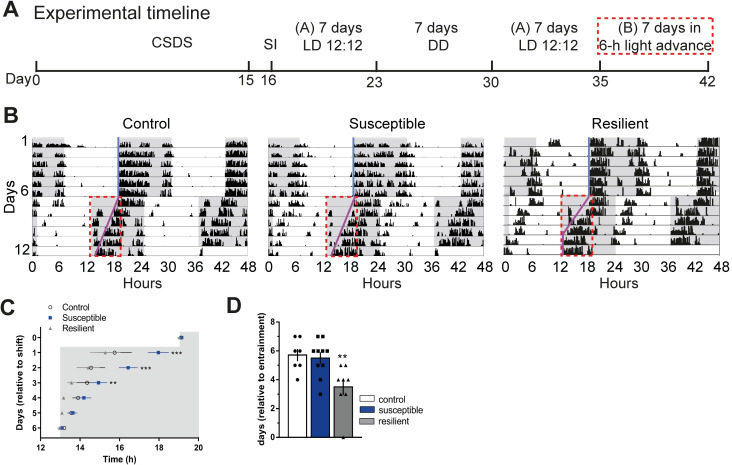
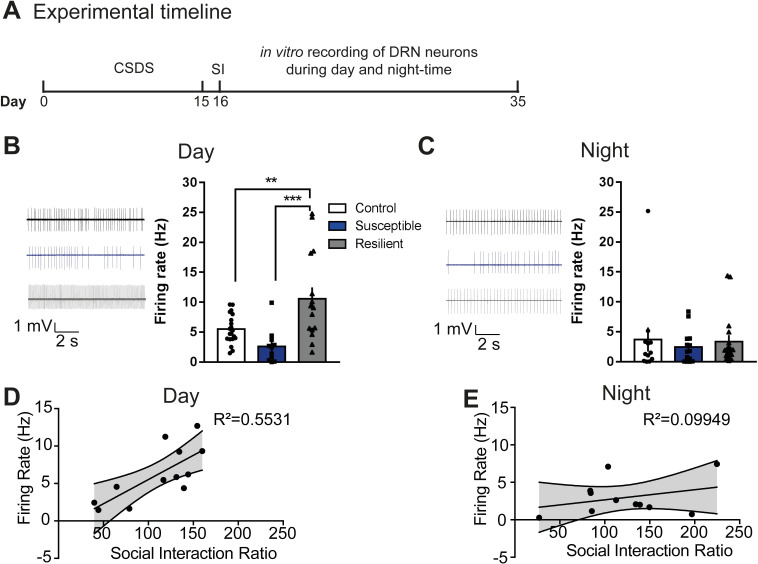
Daily rhythms are disrupted in patients with mood disorders. The lateral habenula (LHb) and dorsal raphe nucleus (DRN) contribute to circadian timekeeping and regulate mood. Thus, pathophysiology in these nuclei may be responsible for aberrations in daily rhythms during mood disorders. Using the 15-day chronic social defeat stress (CSDS) paradigm and in vitro slice electrophysiology, we measured the effects of stress on diurnal rhythms in firing of LHb cells projecting to the DRN (cellsLHb→DRN) and unlabeled DRN cells. We also performed optogenetic experiments to investigate if increased firing in cellsLHb→DRN during exposure to a weak 7-day social defeat stress (SDS) paradigm induces stress-susceptibility. Last, we investigated whether exposure to CSDS affected the ability of mice to photoentrain to a new light-dark (LD) cycle. The cellsLHb→DRN and unlabeled DRN cells of stress-susceptible mice express greater blunted diurnal firing compared to stress-näive (control) and stress-resilient mice. Daytime optogenetic activation of cellsLHb→DRN during SDS induces stress-susceptibility which shows the direct correlation between increased activity in this circuit and putative mood disorders. Finally, we found that stress-susceptible mice are slower, while stress-resilient mice are faster, at photoentraining to a new LD cycle. Our findings suggest that exposure to strong stressors induces blunted daily rhythms in firing in cellsLHb→DRN, DRN cells and decreases the initial rate of photoentrainment in susceptible-mice. In contrast, resilient-mice may undergo homeostatic adaptations that maintain daily rhythms in firing in cellsLHb→DRN and also show rapid photoentrainment to a new LD cycle.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
