

Bacterial defenses against a natural antibiotic promote collateral resilience to clinical antibiotics
- PMID: 33690640
- PMCID: PMC7946323
- DOI: 10.1371/journal.pbio.3001093
Bacterial defenses against a natural antibiotic promote collateral resilience to clinical antibiotics
Abstract
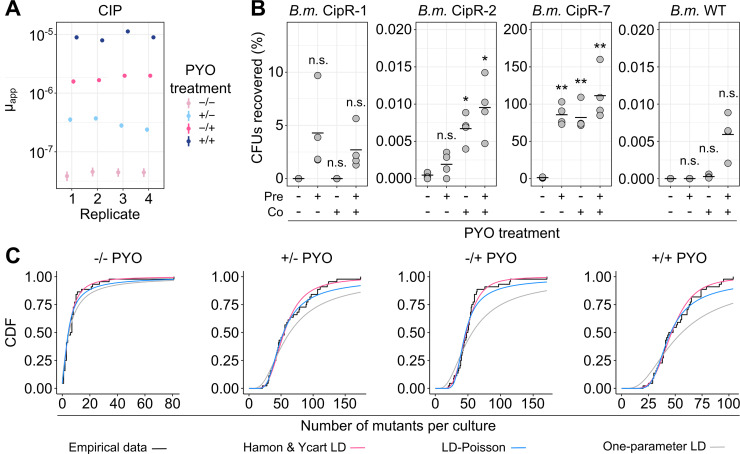
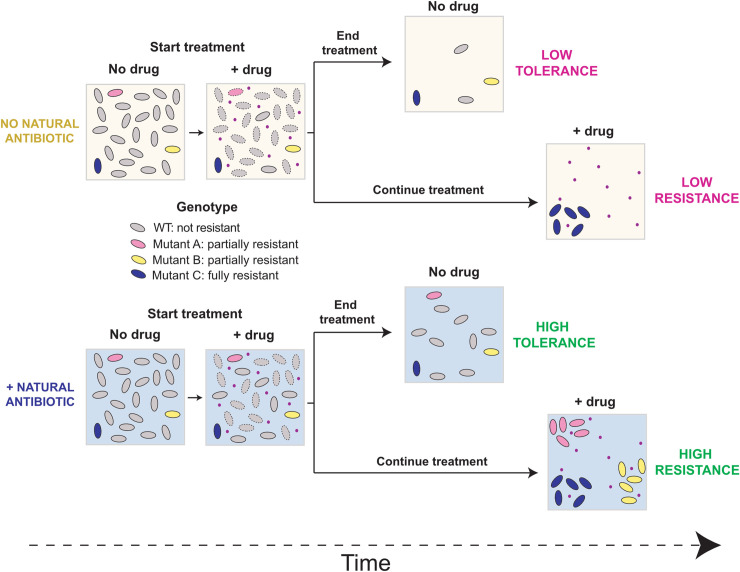
Bacterial opportunistic human pathogens frequently exhibit intrinsic antibiotic tolerance and resistance, resulting in infections that can be nearly impossible to eradicate. We asked whether this recalcitrance could be driven by these organisms' evolutionary history as environmental microbes that engage in chemical warfare. Using Pseudomonas aeruginosa as a model, we demonstrate that the self-produced antibiotic pyocyanin (PYO) activates defenses that confer collateral tolerance specifically to structurally similar synthetic clinical antibiotics. Non-PYO-producing opportunistic pathogens, such as members of the Burkholderia cepacia complex, likewise display elevated antibiotic tolerance when cocultured with PYO-producing strains. Furthermore, by widening the population bottleneck that occurs during antibiotic selection and promoting the establishment of a more diverse range of mutant lineages, PYO increases apparent rates of mutation to antibiotic resistance to a degree that can rival clinically relevant hypermutator strains. Together, these results reveal an overlooked mechanism by which opportunistic pathogens that produce natural toxins can dramatically modulate the efficacy of clinical antibiotics and the evolution of antibiotic resistance, both for themselves and other members of clinically relevant polymicrobial communities.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
