

Regulation of translation by methylation multiplicity of 18S rRNA
- PMID: 33691096
- PMCID: PMC8063911
- DOI: 10.1016/j.celrep.2021.108825
Regulation of translation by methylation multiplicity of 18S rRNA
Abstract
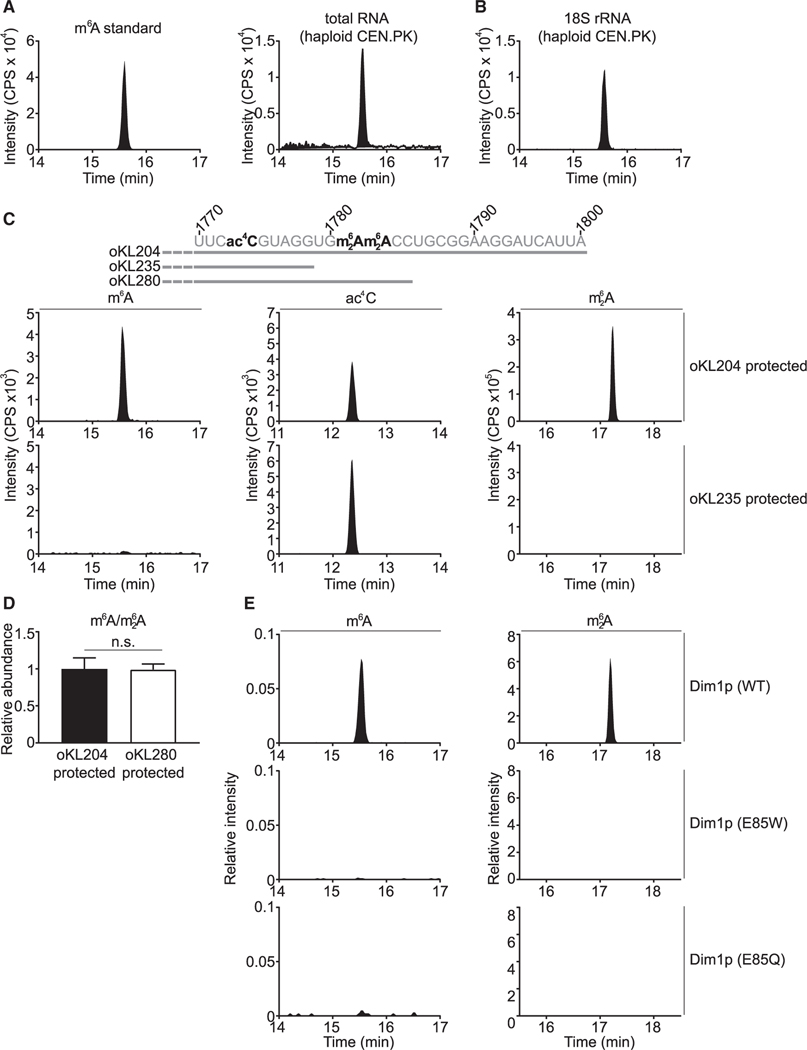
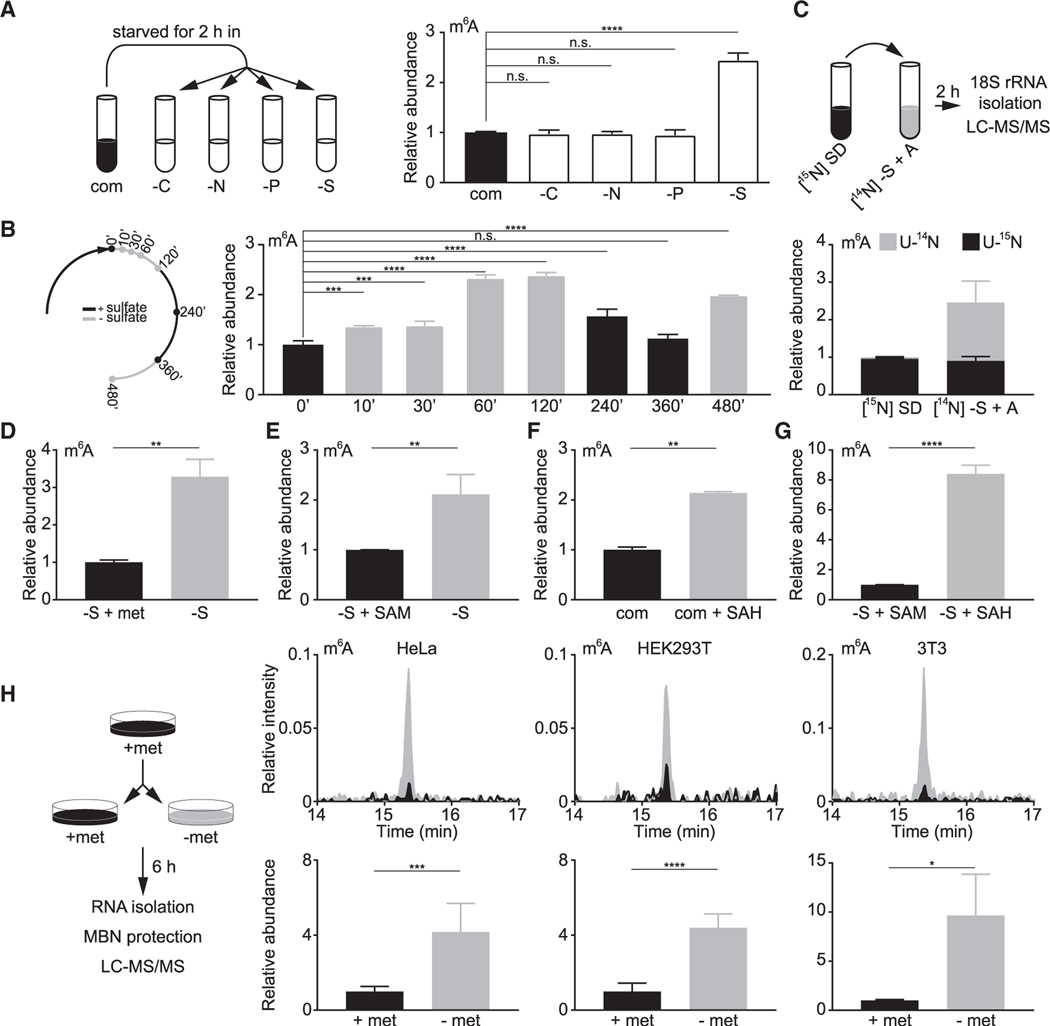
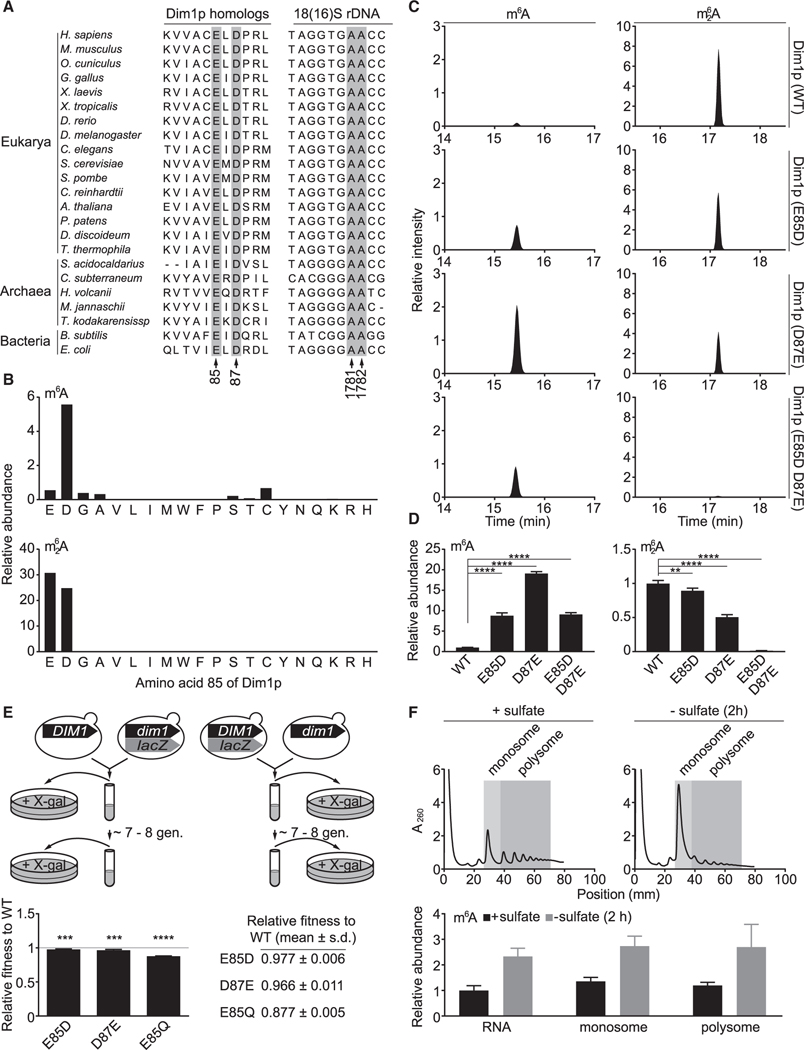
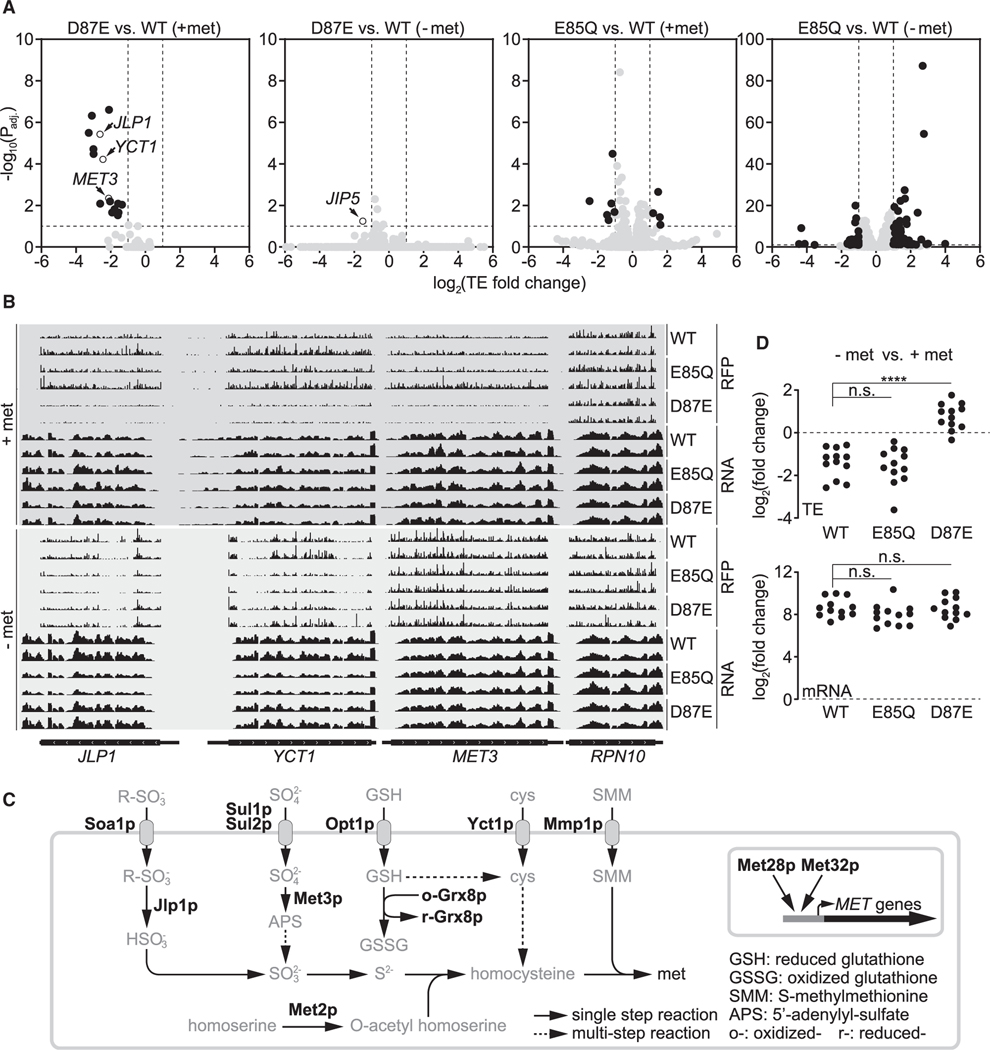
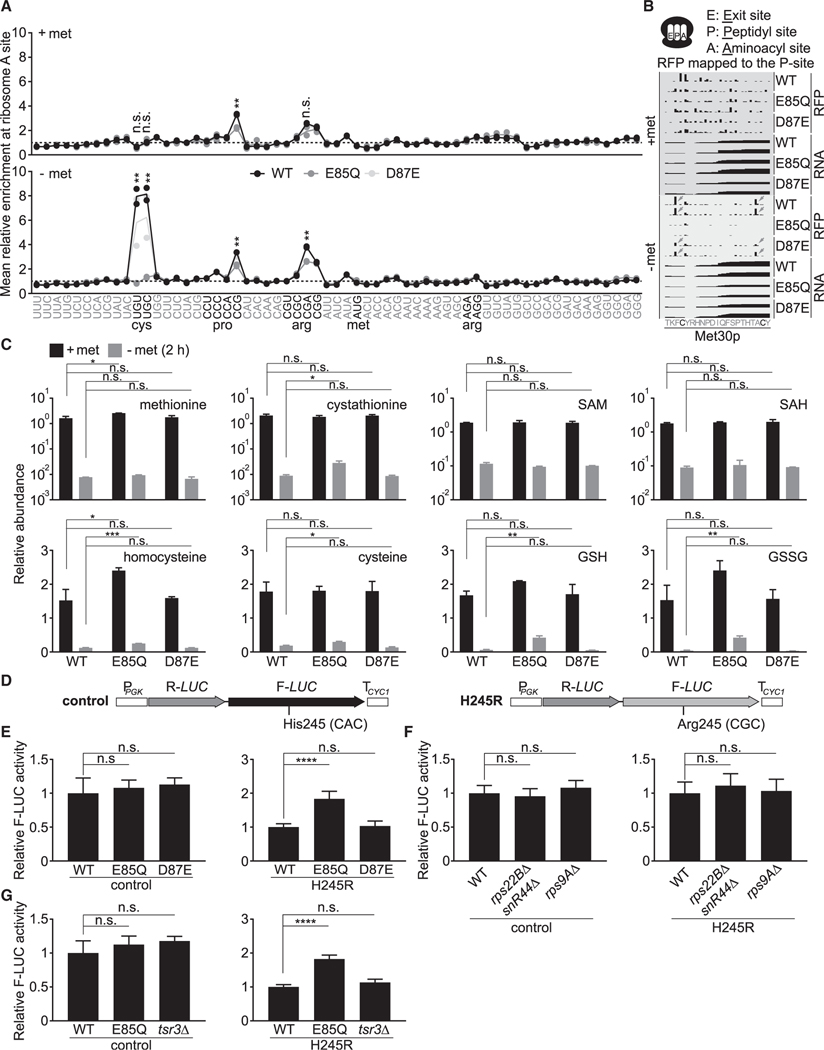
N6-methyladenosine (m6A) is a conserved ribonucleoside modification that regulates many facets of RNA metabolism. Using quantitative mass spectrometry, we find that the universally conserved tandem adenosines at the 3' end of 18S rRNA, thought to be constitutively di-methylated (m62A), are also mono-methylated (m6A). Although present at substoichiometric amounts, m6A at these positions increases significantly in response to sulfur starvation in yeast cells and mammalian cell lines. Combining yeast genetics and ribosome profiling, we provide evidence to suggest that m6A-bearing ribosomes carry out translation distinctly from m62A-bearing ribosomes, featuring a striking specificity for sulfur metabolism genes. Our work thus reveals methylation multiplicity as a mechanism to regulate translation.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Atkinson D (1977). Cellular energy metabolism and its regulation (Academic Press; ), p. 75.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
