

A trans-complementation system for SARS-CoV-2 recapitulates authentic viral replication without virulence
- PMID: 33691138
- PMCID: PMC7901297
- DOI: 10.1016/j.cell.2021.02.044
A trans-complementation system for SARS-CoV-2 recapitulates authentic viral replication without virulence
Abstract
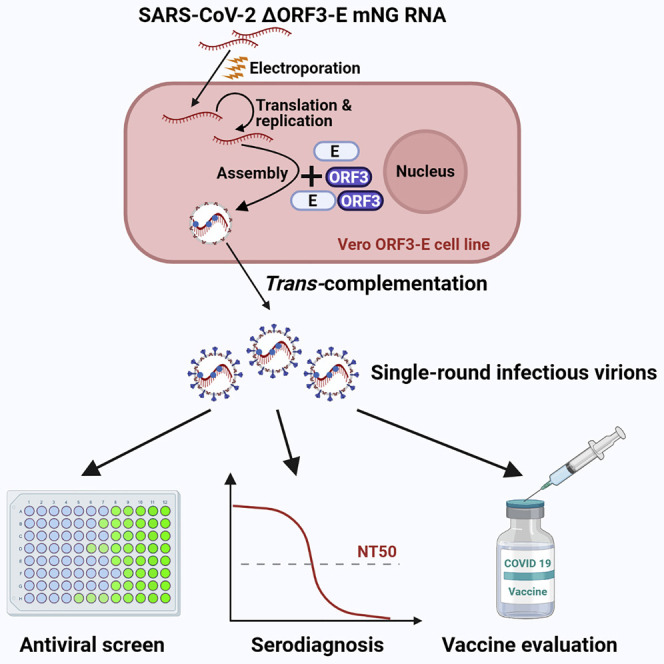
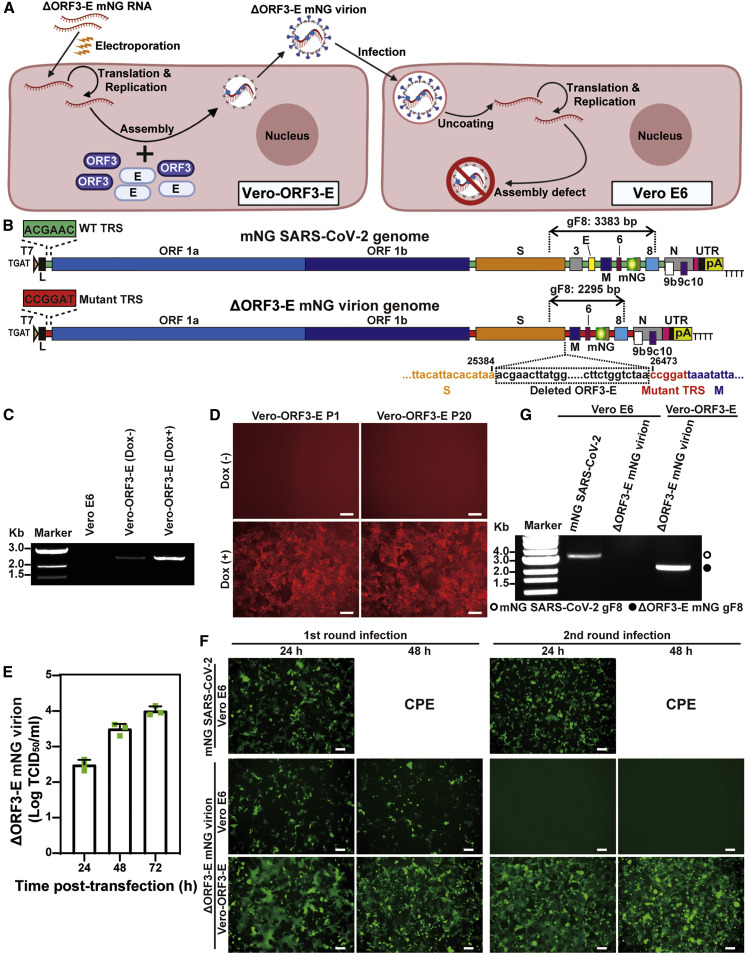
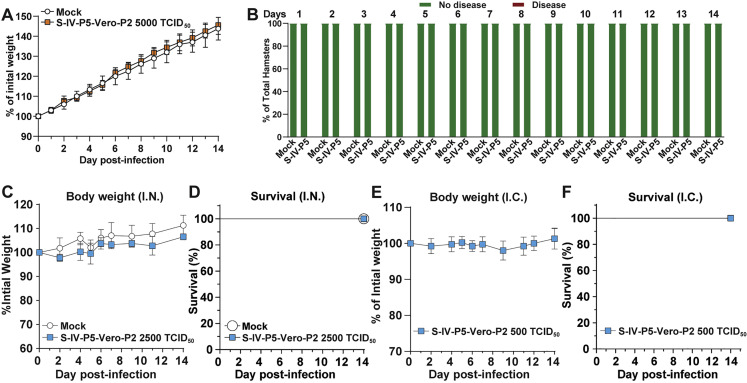
The biosafety level 3 (BSL-3) requirement to culture severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is a bottleneck for research. Here, we report a trans-complementation system that produces single-round infectious SARS-CoV-2 that recapitulates authentic viral replication. We demonstrate that the single-round infectious SARS-CoV-2 can be used at BSL-2 laboratories for high-throughput neutralization and antiviral testing. The trans-complementation system consists of two components: a genomic viral RNA containing ORF3 and envelope gene deletions, as well as mutated transcriptional regulator sequences, and a producer cell line expressing the two deleted genes. Trans-complementation of the two components generates virions that can infect naive cells for only one round but does not produce wild-type SARS-CoV-2. Hamsters and K18-hACE2 transgenic mice inoculated with the complementation-derived virions exhibited no detectable disease, even after intracranial inoculation with the highest possible dose. Thus, the trans-complementation platform can be safely used at BSL-2 laboratories for research and countermeasure development.
Keywords: COVID-19; SARS-CoV-2; antiviral; coronavirus; diagnosis; vaccine.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests X.Z., X.X., and P.-Y.S. have filed a patent on the trans-complementation system of SARS-CoV-2. M.S.D. is a consultant for Inbios, Vir Biotechnology, NGM Biopharmaceuticals, and Carnival Corporation and on the Scientific Advisory Boards of Moderna and Immunome. The Diamond laboratory has received unrelated funding support in sponsored research agreements from Moderna, Vir Biotechnology, and Emergent BioSolutions.
Figures
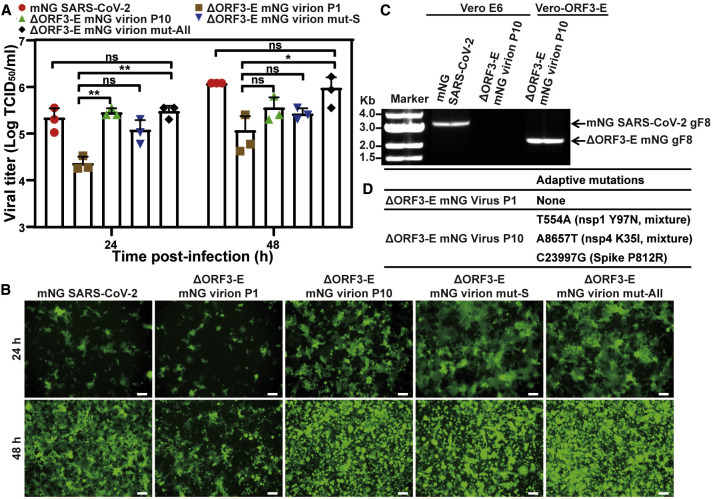
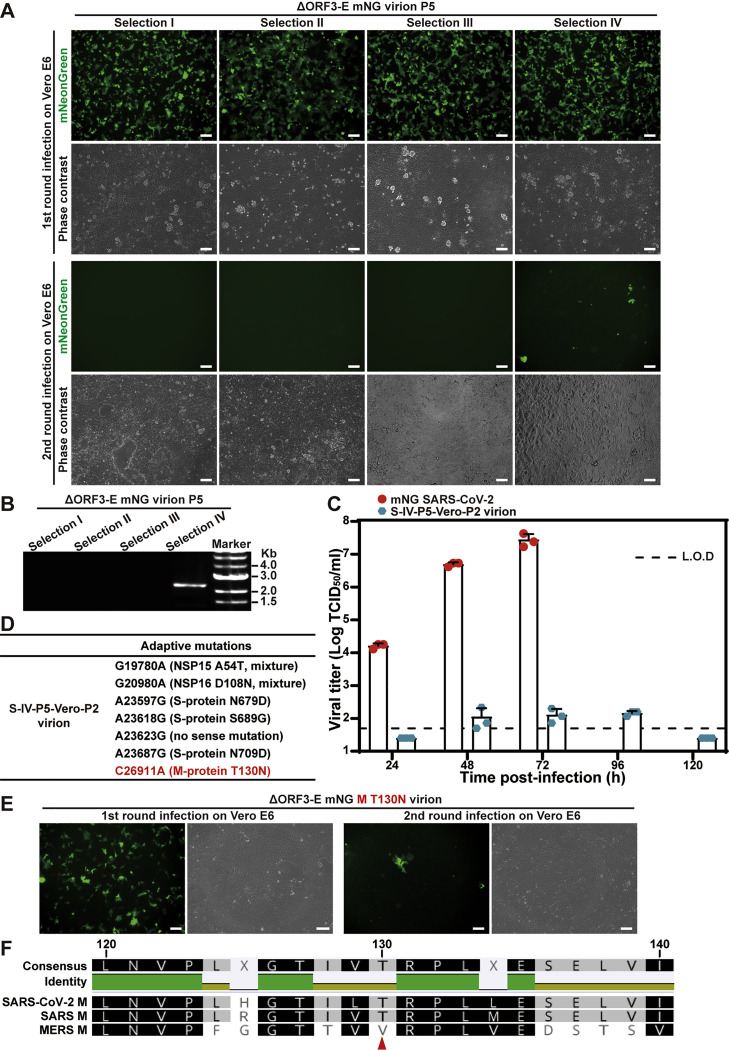
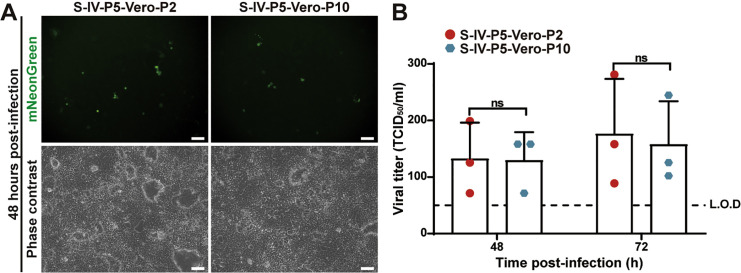
Update of
-
A trans -complementation system for SARS-CoV-2.bioRxiv [Preprint]. 2021 Jan 19:2021.01.16.426970. doi: 10.1101/2021.01.16.426970. bioRxiv. 2021. Update in: Cell. 2021 Apr 15;184(8):2229-2238.e13. doi: 10.1016/j.cell.2021.02.044. PMID: 33501436 Free PMC article. Updated. Preprint.
References
-
- Castaño-Rodriguez C., Honrubia J.M., Gutiérrez-Álvarez J., DeDiego M.L., Nieto-Torres J.L., Jimenez-Guardeño J.M., Regla-Nava J.A., Fernandez-Delgado R., Verdia-Báguena C., Queralt-Martín M., et al. Role of Severe Acute Respiratory Syndrome Coronavirus Viroporins E, 3a, and 8a in Replication and Pathogenesis. MBio. 2018;9:e02325-17. - PMC - PubMed
-
- Chan J.F., Zhang A.J., Yuan S., Poon V.K., Chan C.C., Lee A.C., Chan W.M., Fan Z., Tsoi H.W., Wen L., et al. Simulation of the clinical and pathological manifestations of Coronavirus Disease 2019 (COVID-19) in golden Syrian hamster model: implications for disease pathogenesis and transmissibility. Clin. Infect. Dis. 2020;71:2428–2446. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
