

Deletion of Kcnj16 in Mice Does Not Alter Auditory Function
- PMID: 33693002
- PMCID: PMC7937937
- DOI: 10.3389/fcell.2021.630361
Deletion of Kcnj16 in Mice Does Not Alter Auditory Function
Abstract
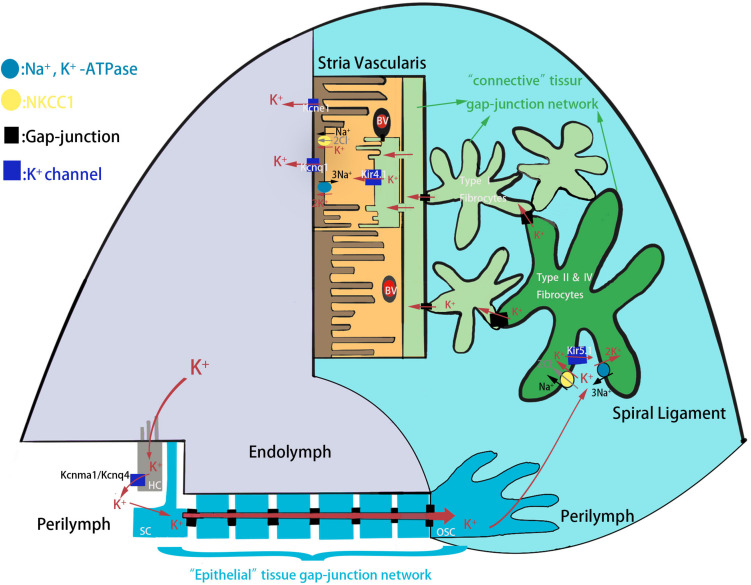
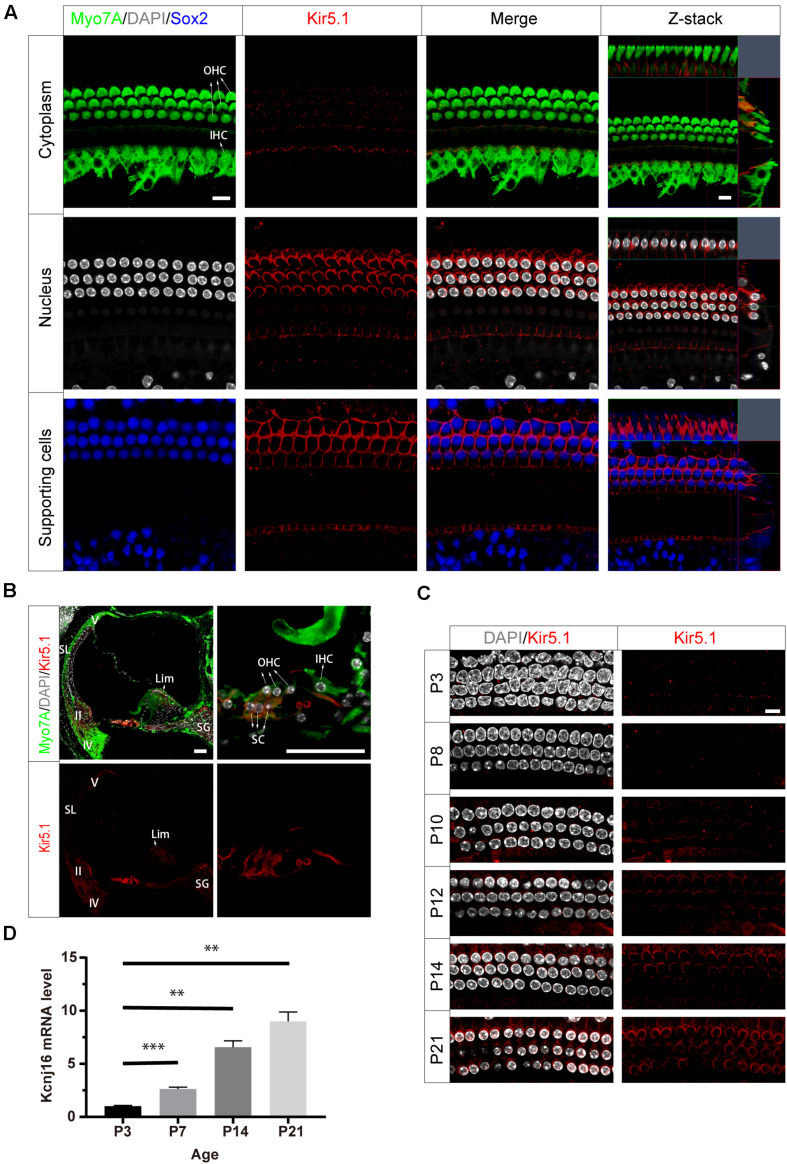
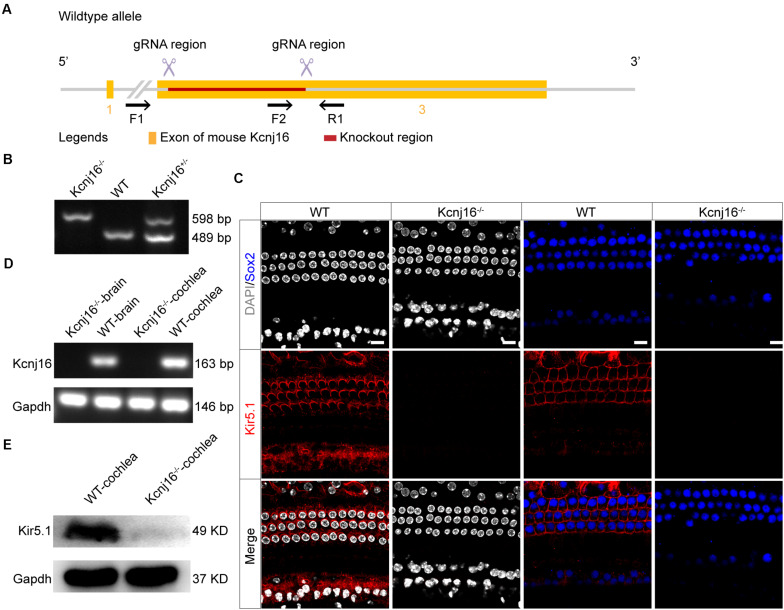
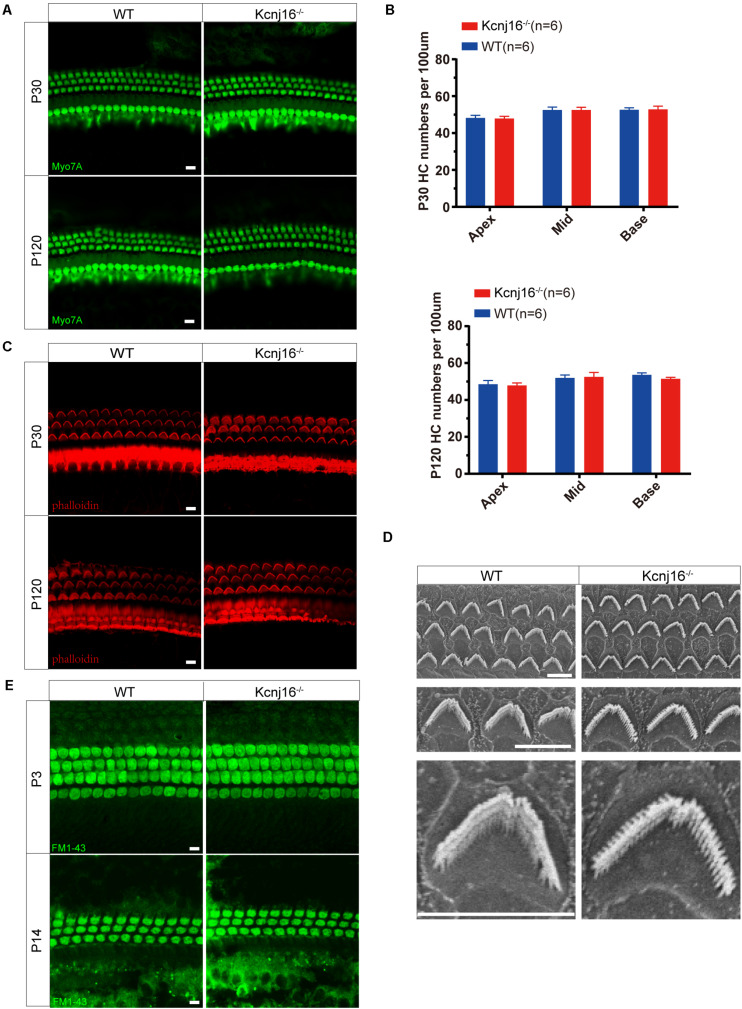
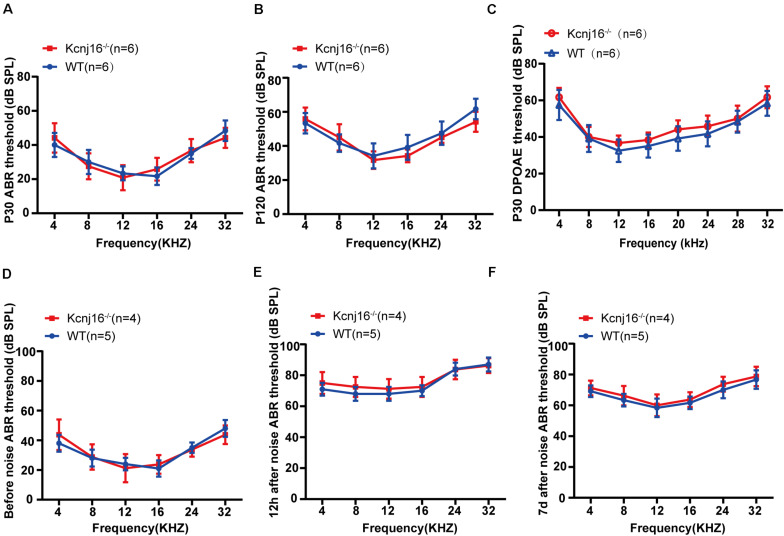
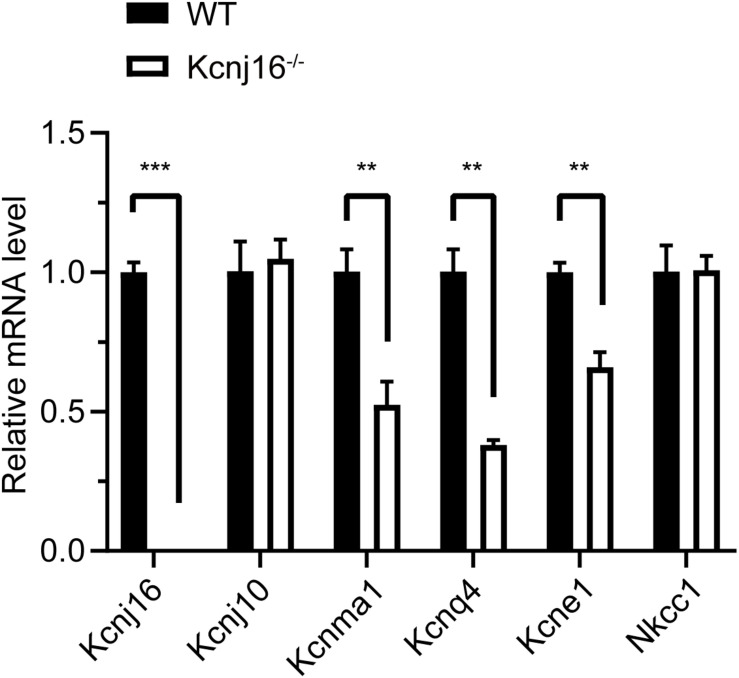
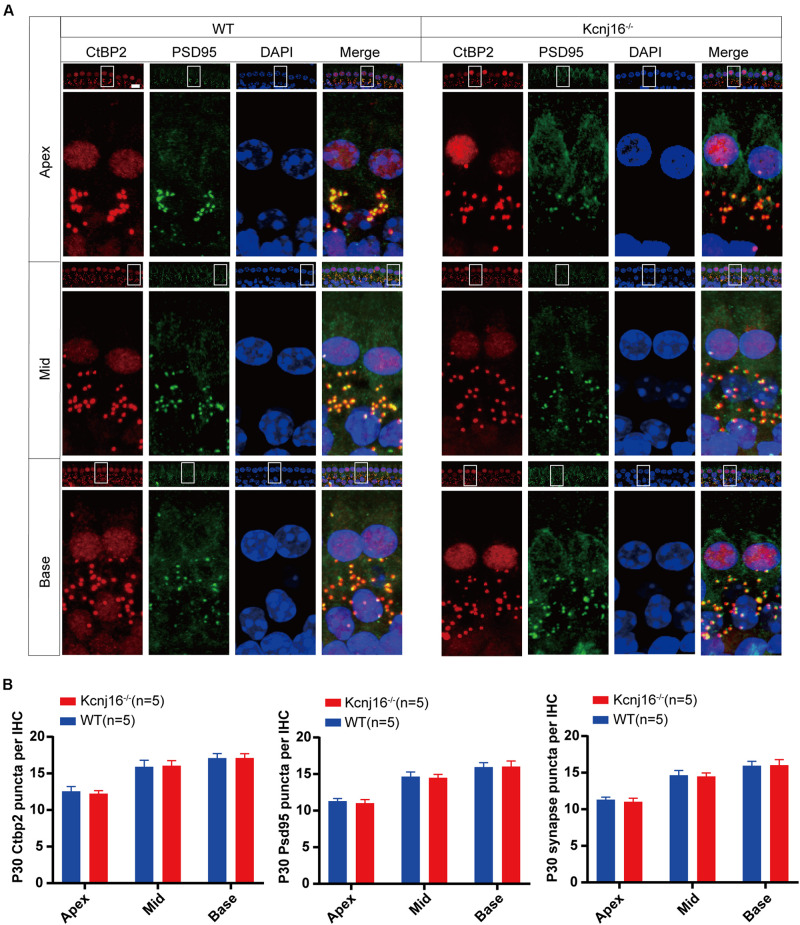
Endolymphatic potential (EP) is the main driving force behind the sensory transduction of hearing, and K+ is the main charge carrier. Kir5.1 is a K+ transporter that plays a significant role in maintaining EP homeostasis, but the expression pattern and role of Kir5.1 (which is encoded by the Kcnj16 gene) in the mouse auditory system has remained unclear. In this study, we found that Kir5.1 was expressed in the mouse cochlea. We checked the inner ear morphology and measured auditory function in Kcnj16 -/- mice and found that loss of Kcnj16 did not appear to affect the development of hair cells. There was no significant difference in auditory function between Kcnj16 -/- mice and wild-type littermates, although the expression of Kcnma1, Kcnq4, and Kcne1 were significantly decreased in the Kcnj16 -/- mice. Additionally, no significant differences were found in the number or distribution of ribbon synapses between the Kcnj16 -/- and wild-type mice. In summary, our results suggest that the Kcnj16 gene is not essential for auditory function in mice.
Keywords: Kir5.1; cochlea; endolymphatic potential; hair cell; hearing loss.
Copyright © 2021 Lv, Fu, Li, Hong, Li, Lin, Xun, Fang, Weng, Yue, Li, Guan, Li, Huang and Chai.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases