

The multi-scale architecture of mammalian sperm flagella and implications for ciliary motility
- PMID: 33694216
- PMCID: PMC8013824
- DOI: 10.15252/embj.2020107410
The multi-scale architecture of mammalian sperm flagella and implications for ciliary motility
Abstract
Motile cilia are molecular machines used by a myriad of eukaryotic cells to swim through fluid environments. However, available molecular structures represent only a handful of cell types, limiting our understanding of how cilia are modified to support motility in diverse media. Here, we use cryo-focused ion beam milling-enabled cryo-electron tomography to image sperm flagella from three mammalian species. We resolve in-cell structures of centrioles, axonemal doublets, central pair apparatus, and endpiece singlets, revealing novel protofilament-bridging microtubule inner proteins throughout the flagellum. We present native structures of the flagellar base, which is crucial for shaping the flagellar beat. We show that outer dense fibers are directly coupled to microtubule doublets in the principal piece but not in the midpiece. Thus, mammalian sperm flagella are ornamented across scales, from protofilament-bracing structures reinforcing microtubules at the nano-scale to accessory structures that impose micron-scale asymmetries on the entire assembly. Our structures provide vital foundations for linking molecular structure to ciliary motility and evolution.
Keywords: centrioles; cryo-FIB milling; cryo-electron tomography; motile cilia; sperm.
© 2021 The Authors. Published under the terms of the CC BY NC ND 4.0 license.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
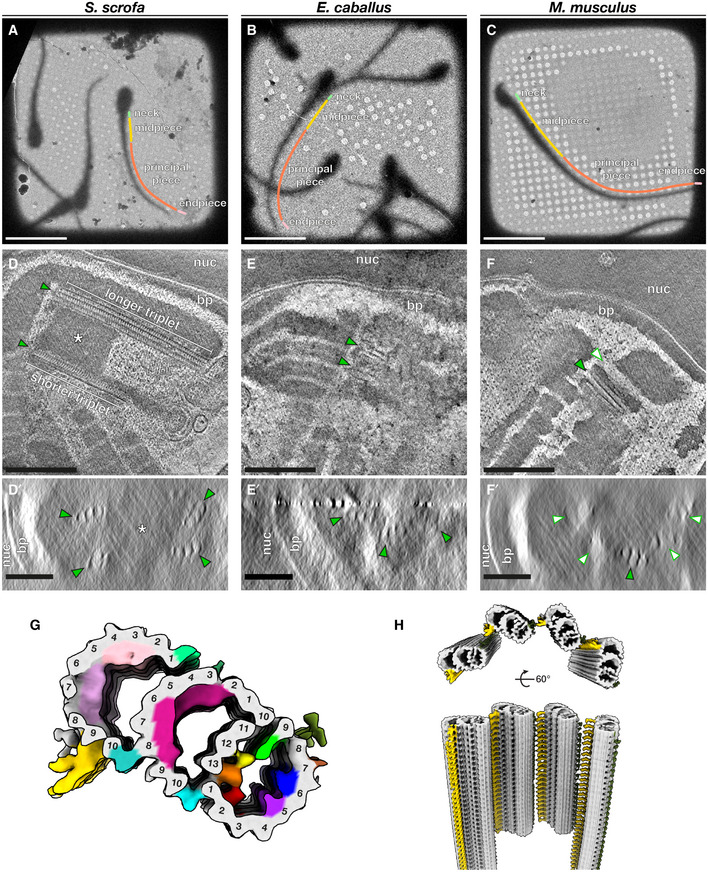
- A–C
Low‐magnification cryo‐EM projection images of pig (A), horse (B), and mouse (C) sperm. Different regions of the flagellum discussed in this paper are annotated as follows: green—neck, yellow—midpiece, coral—principal piece, pink—endpiece.
- D–F
Tomographic slices through cryo‐FIB‐milled lamellae of pig (D), horse (E), and mouse (F) sperm. Transverse slices (D’–F’) show complete triplets in the pig (D’) and the horse (E’), but not in the mouse (F’). Complete triplets are indicated by green arrowheads with black outlines, while degenerated triplets are indicated by white arrowheads with green outlines. Note the electron‐dense material in the lumen of the pig sperm proximal centriole that is continuous with the connecting piece (asterisks in D and D’).
- G
In situ structure of triplet microtubules from the proximal region of the pig sperm PC with the tubulin backbone in gray and microtubule inner protein densities colored individually. A‐tubule MIPs are colored: MIP1—green, MIP2—yellow, MIP3—orange, MIP4—red, MIP5—purple, MIP6—blue. B‐tubule MIPs are colored: MIP7—magenta. C‐tubule MIPs are colored: MIP8—light pink, MIP9—pink. The A‐C linker is colored gold and the putative A‐link is colored olive green.
- H
Reconstruction of the proximal region of the pig sperm PC generated by plotting the average back into the original particle positions and orientations in the tomogram. This plotback only contains four triplets as only part of the centriole was captured in the lamella. The A–C linker is colored in gold and the putative A‐link in olive green.

PC triplets have unequal lengths (top panel), and shorter triplets are grouped on one side, giving the PC dorsoventral asymmetry (bottom panel).
Many of the microtubule inner proteins (MIPs) in the pig sperm PC are not found in other mammalian centriole structures.
Details of the MIP densities in the pig sperm PC.
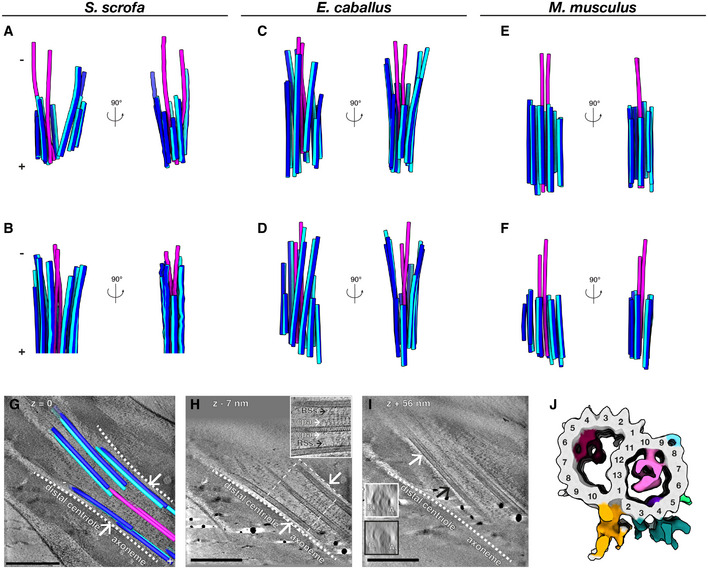
- A–F
Microtubules in the DC of pig (A, B), horse (C, D), and mouse (E, F) sperm traced from Volta phase plate cryo‐tomograms of intact sperm. Doublets are colored blue (A‐tubule in light blue, B‐tubule in dark blue), while singlets are pink.
- G–I
Tomographic slices through cryo‐FIB‐milled lamellae of the DC‐to‐axoneme transition in pig sperm show how the change in geometry (G, white arrows and white dashed lines) coincides with the appearance of axoneme accessory structures (H, white arrows) and with density in the A‐tubule (I, compare insets in white and black boxes). In (I), the white and black arrows indicate where the cross‐sections in white and black boxes were taken from.
- J
In situ structure of the pig sperm DC microtubule doublet with the tubulin backbone in gray and microtubule inner protein densities colored individually. The gold and turquoise densities on the luminal side of the doublet are consistent with the positions of parts of the inner scaffold. This structure represents the DC doublets closest to the axoneme (the area shown in (G–I)).
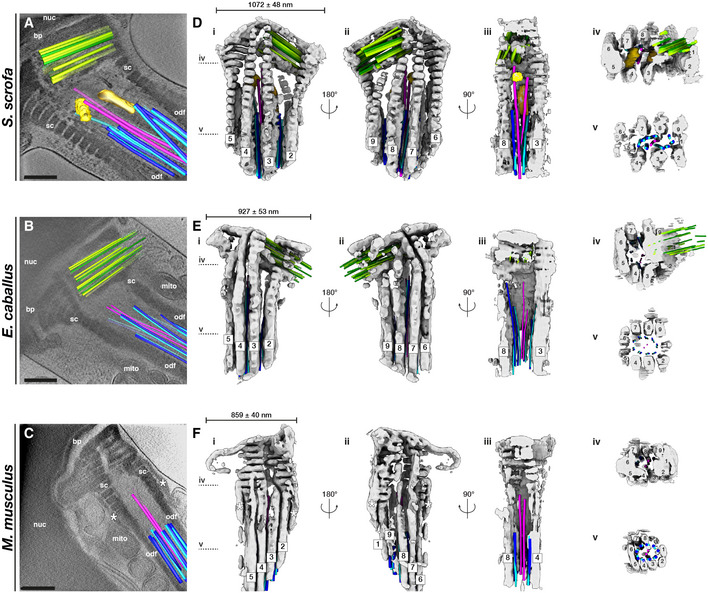
- A–C
Slices through Volta phase plate cryo‐tomograms of the neck region in intact pig (A), horse (B), and mouse (C) sperm. Proximal centriole triplets are shown in green, distal centriole doublets in blue (A‐tubule in light blue, B‐tubule in dark blue) and singlets in pink, and electron‐dense bars in yellow. Note the electron‐dense structures flanking the connecting piece in the mouse (asterisks in C, F).
- D–F
Three‐dimensional architecture of the flagellar base, with the connecting piece in gray, the proximal centriole in green, distal centriole doublets in blue (A‐tubule in light blue, B‐tubule in dark blue) and singlets in pink, and electron‐dense bars in yellow. The connecting piece was segmented semi‐automatically with a neural network, while microtubules were traced manually.
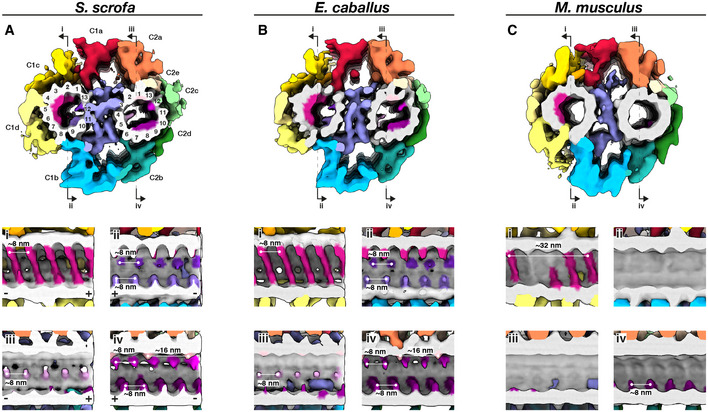
- A–C
Whole‐population in situ structures of the 32‐nm CPA repeat from pig (A), horse (B), and mouse (C) sperm. Panels (i), (ii), (iii), and (iv) are cut‐through views of the lumens of the C1 (i and ii) and C2 (iii and iv) microtubules. Microtubules are shown in gray and microtubule protein densities are colored individually. The projection network is colored according to (Carbajal‐González et al, 2013).

- A–C
In situ structures of the 96‐nm repeat from pig (A), horse (B), and mouse (C) sperm principal pieces.
- D–F
Classification focused on the RS1 barrel revealed two distinct classes of particles, one with (top panels) and one without (middle panels) the structure. Particles with the RS1 barrel are distributed asymmetrically around the axoneme (bottom panels).
- G–I
Microtubule inner proteins in axonemes from pig (G), horse (H), and mouse (I) sperm.
- J–N
Microtubule inner proteins in axonemes from other cell types and organisms. Arrowheads correspond to locations of MIPs that are prominent in mammalian sperm: magenta—helical MIP in the B‐tubule; pink—large A‐tubule MIP; light pink—additional density associated with MIP3a.
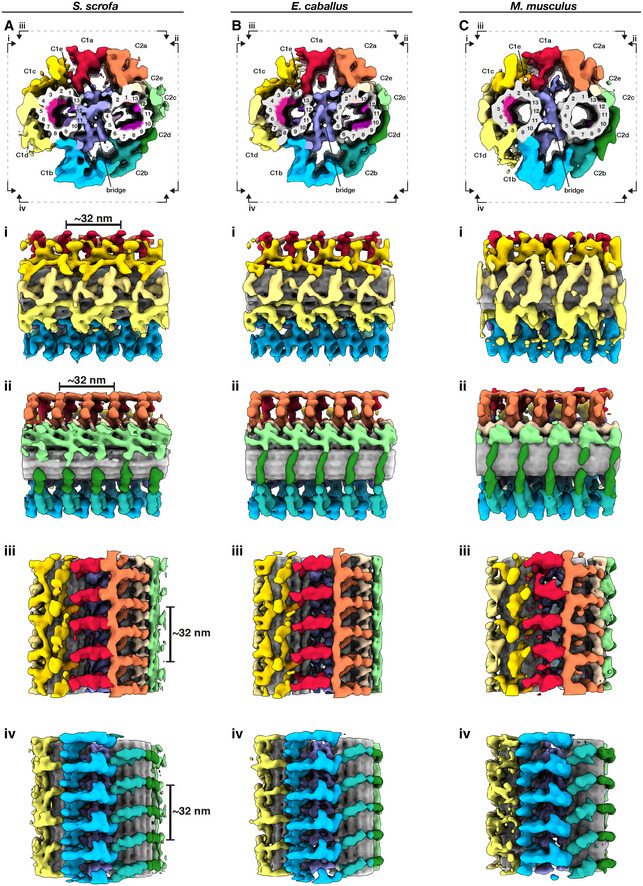
- A–C
Whole‐population in situ structures of the 32‐nm CPA repeat from pig (A), horse (B), and mouse (C) sperm. Individual rotated views show details of C1 projections (i), C2 projections (ii), the interface between C1a and C2a projections (iii), and the interface between C1b and C2b projections (iv).
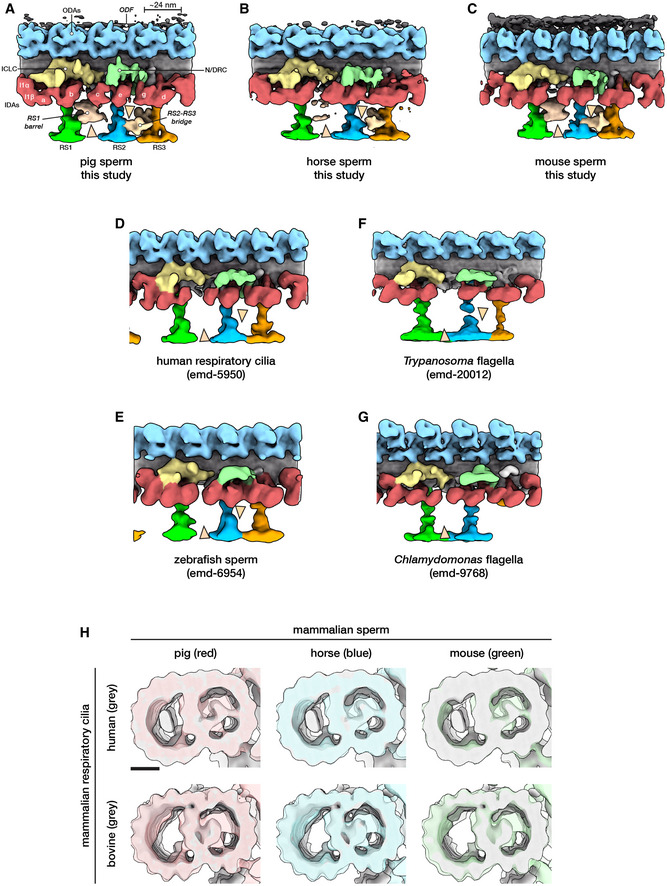
- A–G
Structures of the 96‐nm axonemal repeat from various species in cell types. Arrowheads mark the novel RS1 barrel and RS2‐RS3 bridge so far seen only in mammalian sperm.
- H
A‐tubule MIPs are present in mammalian respiratory cilia, but the A‐MIP in mammalian sperm is larger and makes more extensive connections with the A‐tubule itself. Scale bar: 10 nm.

- A–C
In situ structures of the 96‐nm axonemal repeat from the principal piece of pig (A), horse (B), and mouse (C) sperm after classification focused on the ODF attachment.
- D–L
Axoneme structures with particles from tomograms from (D, G, J) only the midpiece, (E, H, K) only the proximal principal piece, and (F, I, L) only the distal principal piece. Left panels (D–L) show transverse slices through sperm flagella, while right panels (D’–L’) show subtomogram averages of the 96‐nm axoneme repeat from the corresponding regions of the tail. In the left panels, one doublet‐ODF pair is traced to emphasize the gap between the ODF and the doublet in the midpiece. The ODF is traced in gray with a black outline, and the doublet is traced in blue (light blue for the A‐tubule, dark blue for B‐tubule). Doublets 3 and 8 are numbered to aid visualization.
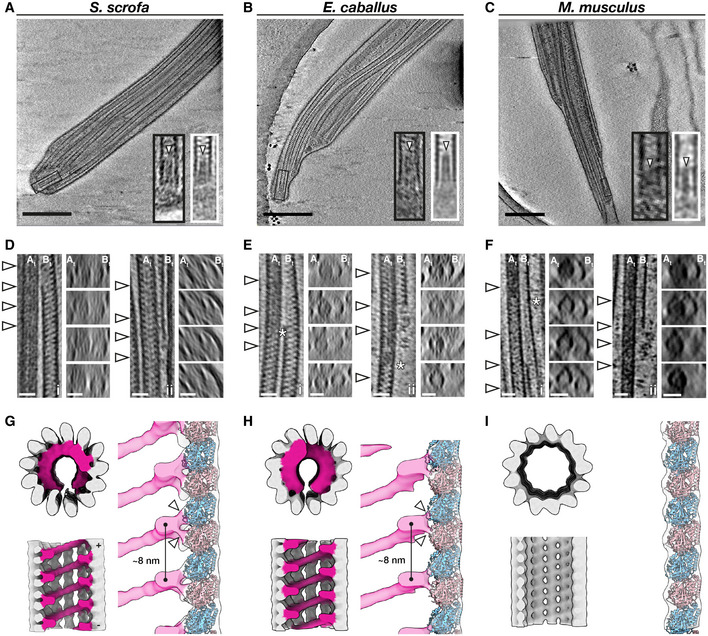
- A–C
Slices through defocus cryo‐tomograms of the endpiece in pig (A), horse (B), and mouse (C) sperm. Insets show digital zooms (black boxes) and subtomogram averages (white boxes) of microtubule tips. Note the presence of a plug (arrowhead) extending into the lumen.
- D–F
Representative examples of the mechanisms by which doublets can transition into singlets. Doublets can split into two complete singlets (panels i), or the B‐tubule can terminate abruptly with the A‐tubule extending as a singlet (panels ii). White arrowheads in the left panels indicate approximate locations of transverse slices shown in the right panels. Asterisks mark approximate locations of the change in A‐tubule luminal density.
- G–I
In situ structures of singlet microtubules from pig (G), horse (H), and mouse (I) sperm endpieces. Note how the MIP binds to two tubulin monomers (white arrowheads).
References
-
- Brokaw CJ (2009) Thinking about flagellar oscillation. Cell Motil Cytoskeleton 66: 425–436 - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
