

UGT84F9 is the major flavonoid UDP-glucuronosyltransferase in Medicago truncatula
- PMID: 33694362
- PMCID: PMC8133618
- DOI: 10.1093/plphys/kiab016
UGT84F9 is the major flavonoid UDP-glucuronosyltransferase in Medicago truncatula
Abstract
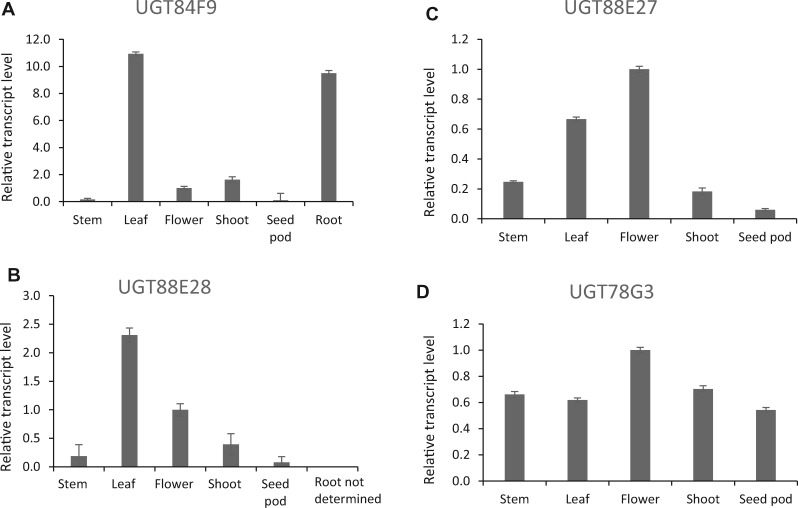
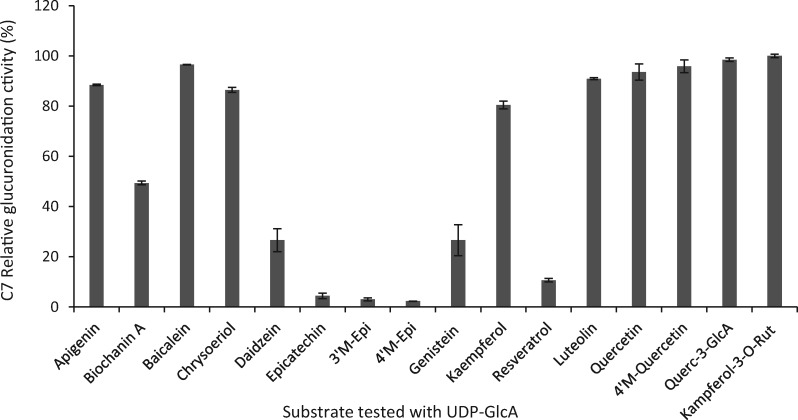
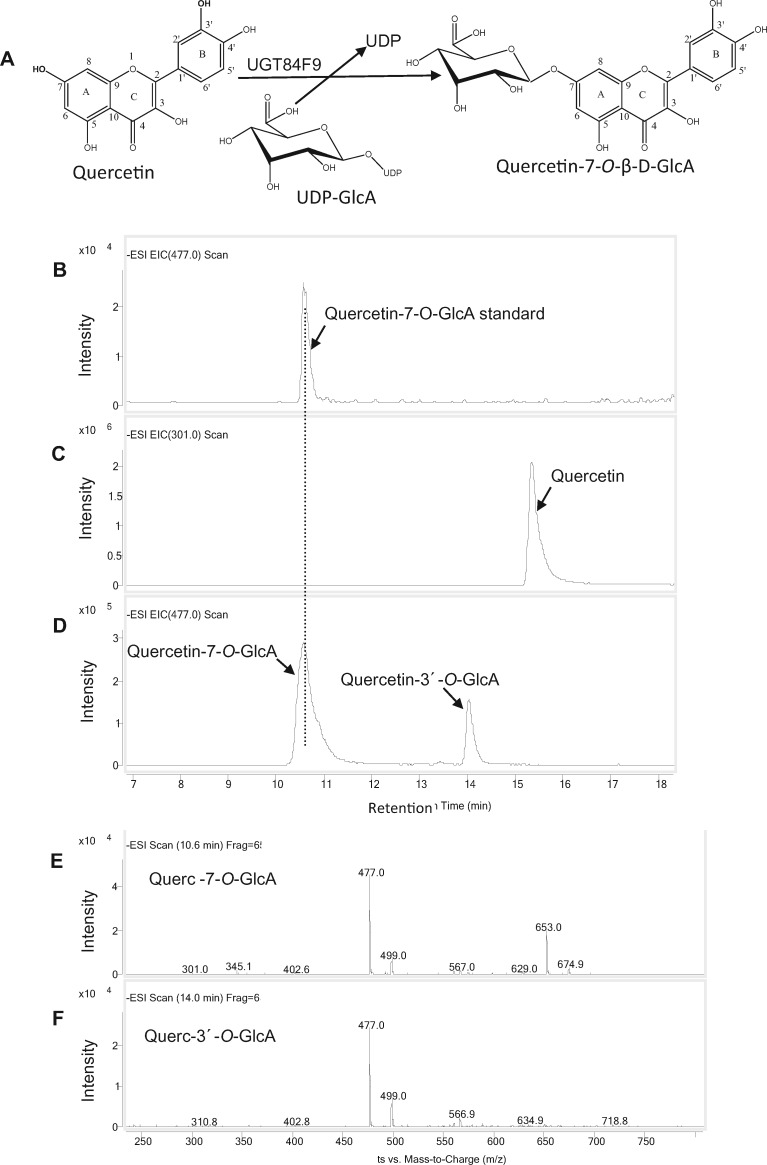
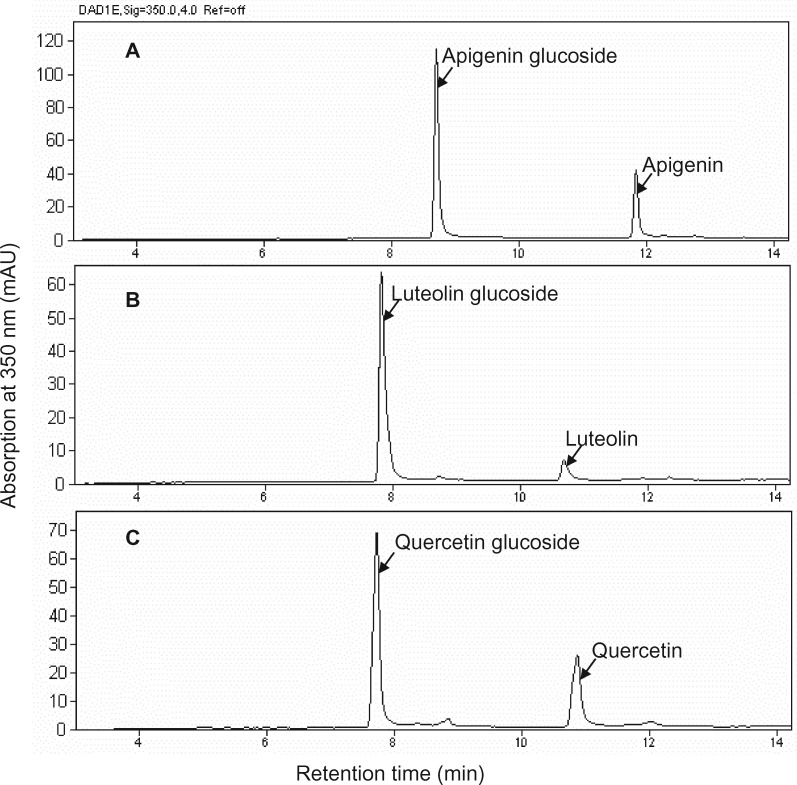
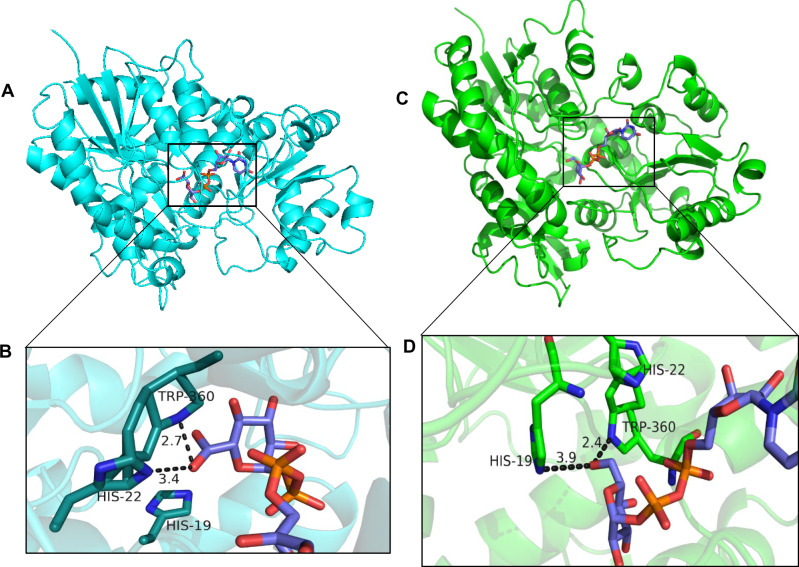
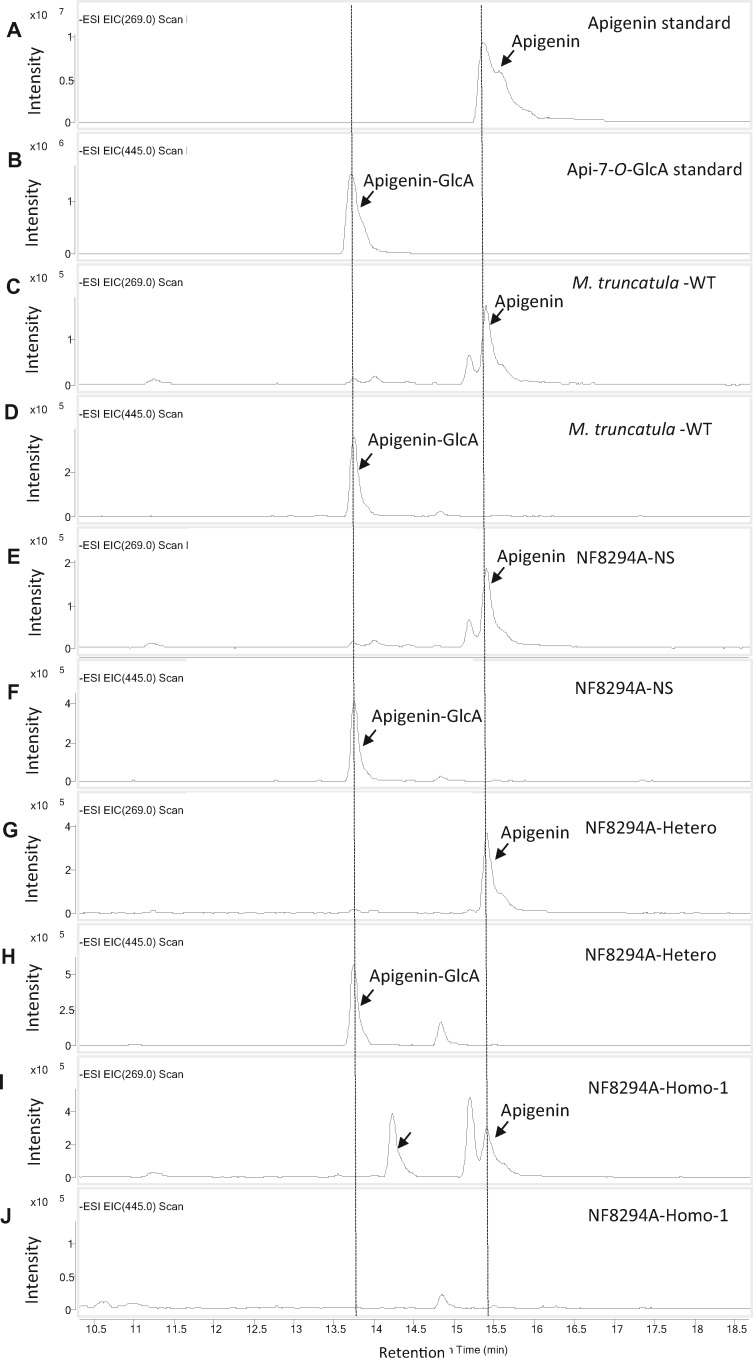
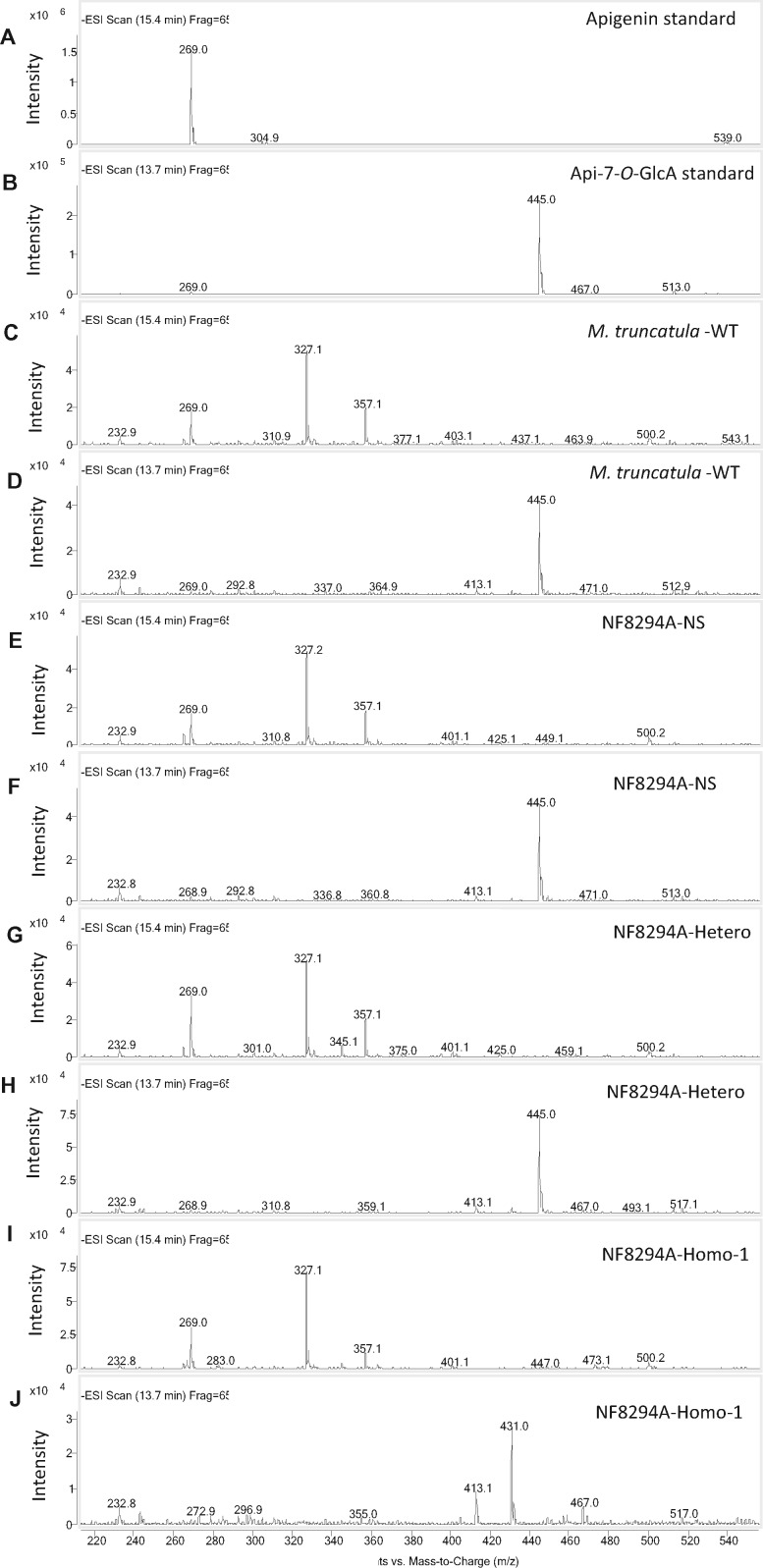
Mammalian phase II metabolism of dietary plant flavonoid compounds generally involves substitution with glucuronic acid. In contrast, flavonoids mainly exist as glucose conjugates in plants, and few plant UDP-glucuronosyltransferase enzymes have been identified to date. In the model legume Medicago truncatula, the major flavonoid compounds in the aerial parts of the plant are glucuronides of the flavones apigenin and luteolin. Here we show that the M. truncatula glycosyltransferase UGT84F9 is a bi-functional glucosyl/glucuronosyl transferase in vitro, with activity against a wide range of flavonoid acceptor molecules including flavones. However, analysis of metabolite profiles in leaves and roots of M. truncatula ugt84f9 loss of function mutants revealed that the enzyme is essential for formation of flavonoid glucuronides, but not most flavonoid glucosides, in planta. We discuss the use of plant UGATs for the semi-synthesis of flavonoid phase II metabolites for clinical studies.
© American Society of Plant Biologists 2021. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Figures


References
-
- Achnine L, Huhman DV, Farag MA, Sumner LW, Blount JW, Dixon RA (2005) Genomics-based selection and functional characterization of triterpene glycosyltransferases from the model legume Medicago truncatula. Plant J 41:875–887 - PubMed
-
- Blount JW, Redan BW, Ferruzzi MG, Reuhs BL, Cooper BR, Harwood JS, Shulaev V, Pasinetti G, Dixon RA (2015) Synthesis and quantitative analysis of plasma-targeted metabolites of catechin and epicatechin. J Agric Food Chem 63:2233–2240 - PubMed
-
- Bowles D, Isayenkova J, Lim E-K, Poppenberger B (2005) Glycosyltransferases: managers of small molecules. Curr Opin Plant Biol 8:254–263 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
