

FUS-dependent liquid-liquid phase separation is important for DNA repair initiation
- PMID: 33704371
- PMCID: PMC7953258
- DOI: 10.1083/jcb.202008030
FUS-dependent liquid-liquid phase separation is important for DNA repair initiation
Abstract
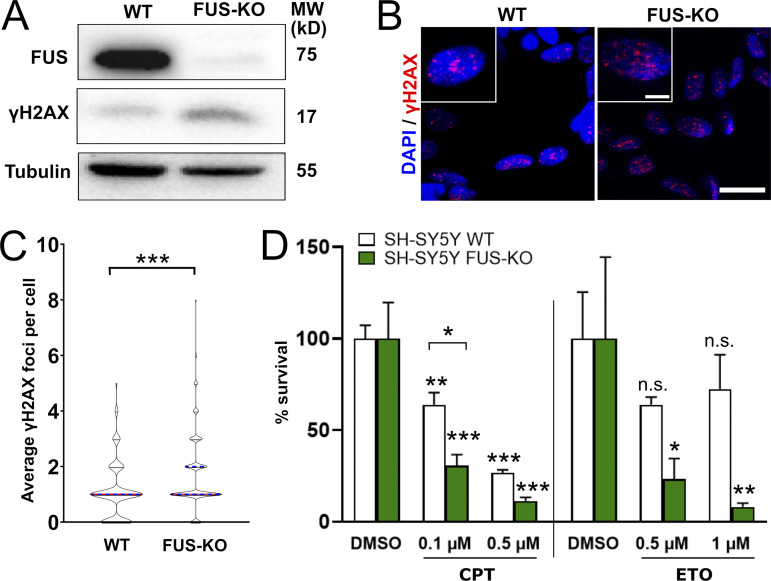
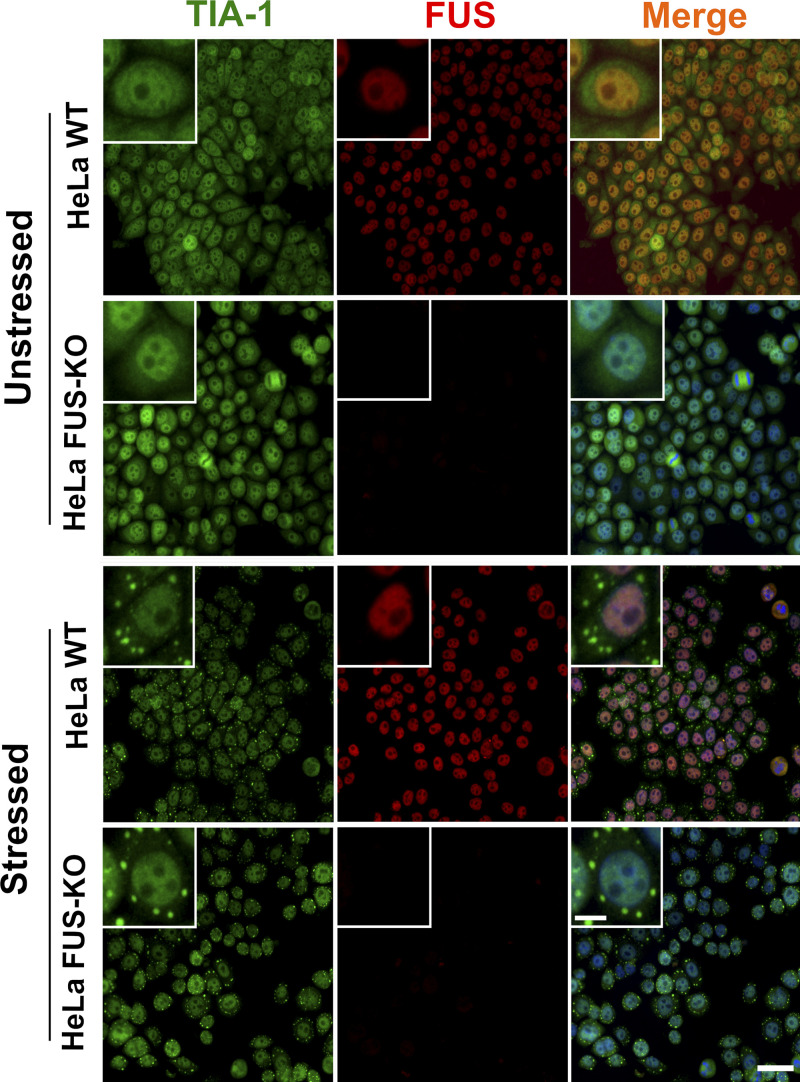
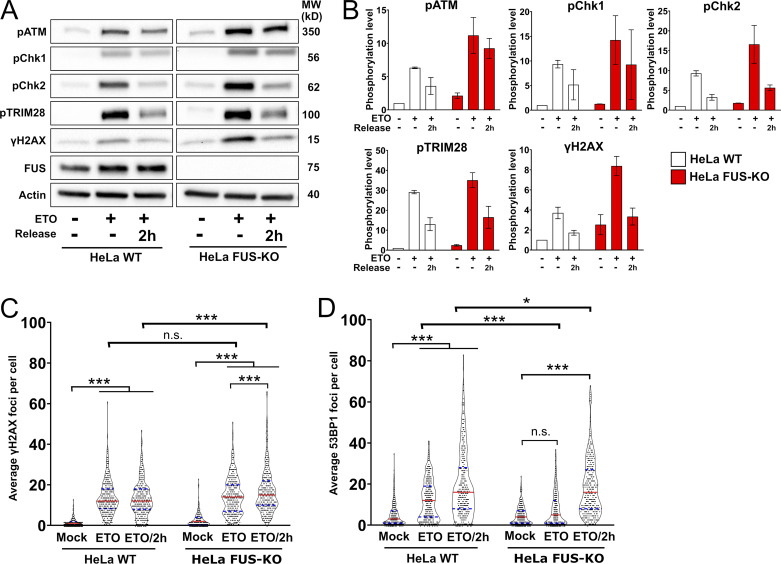
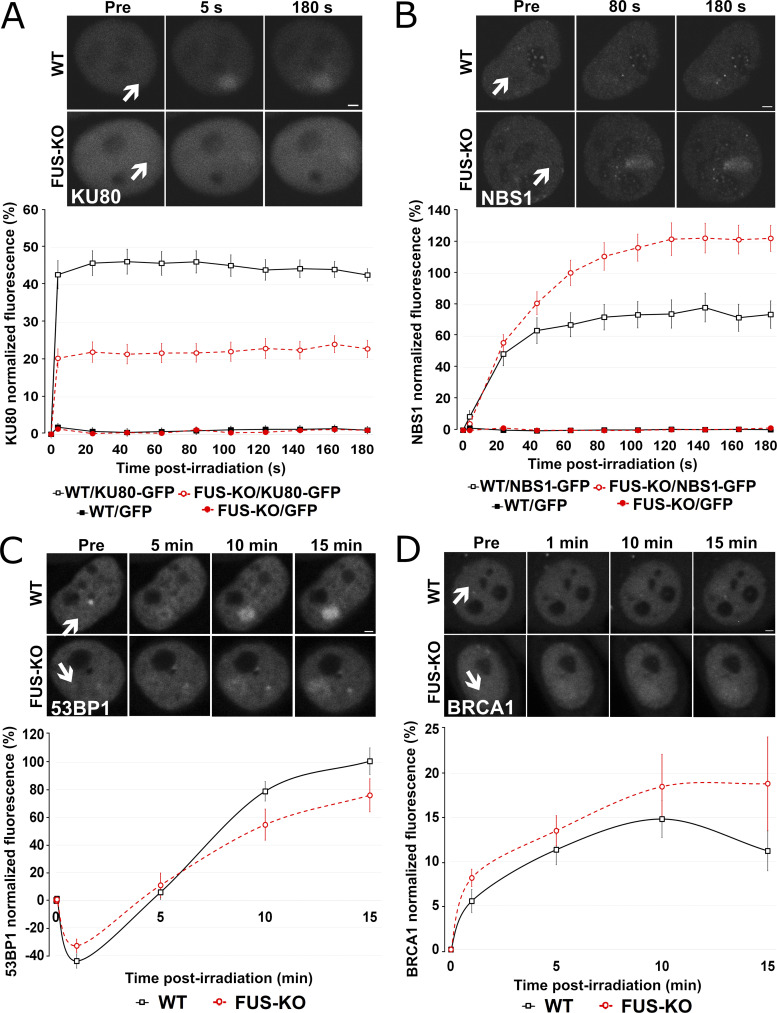
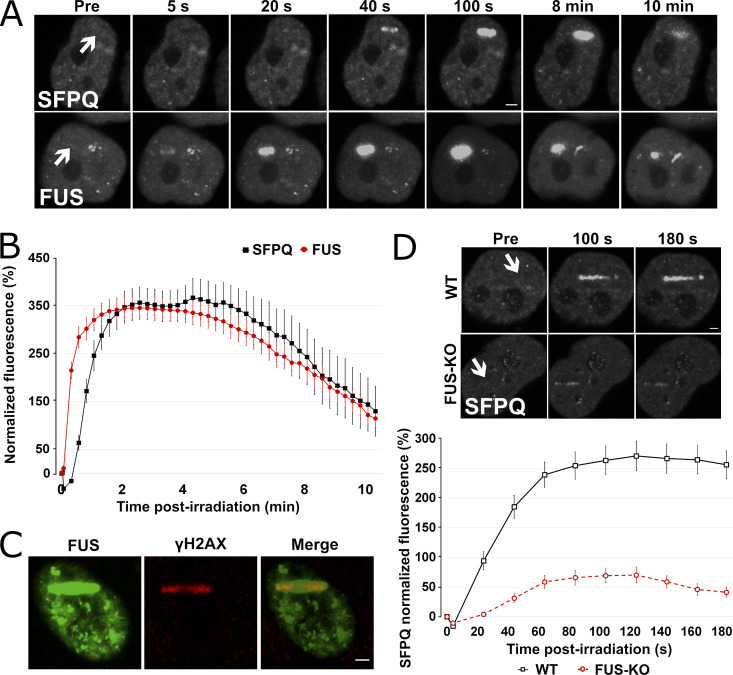
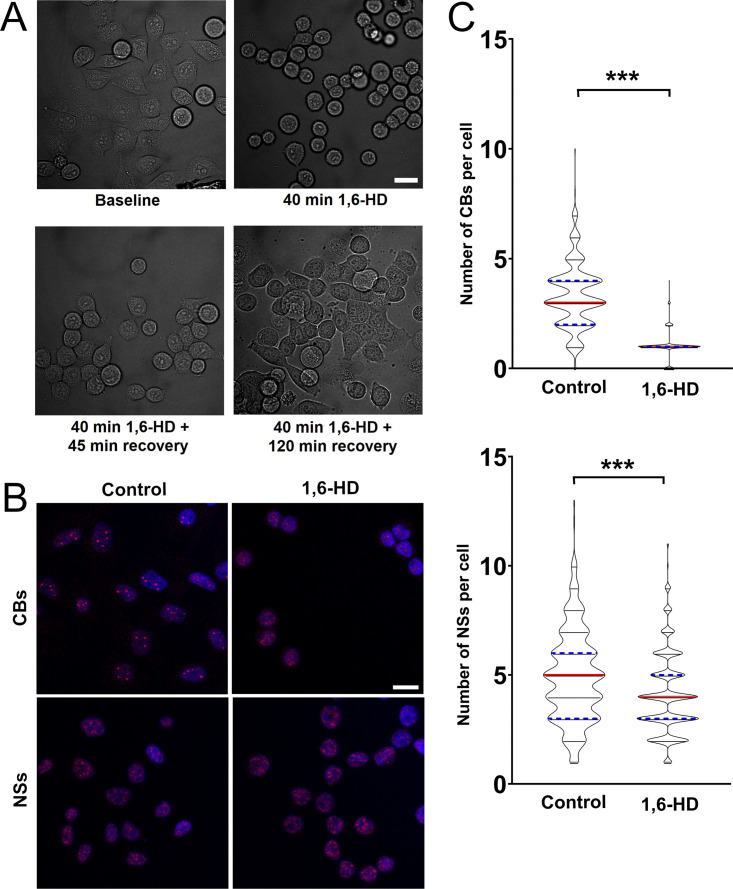
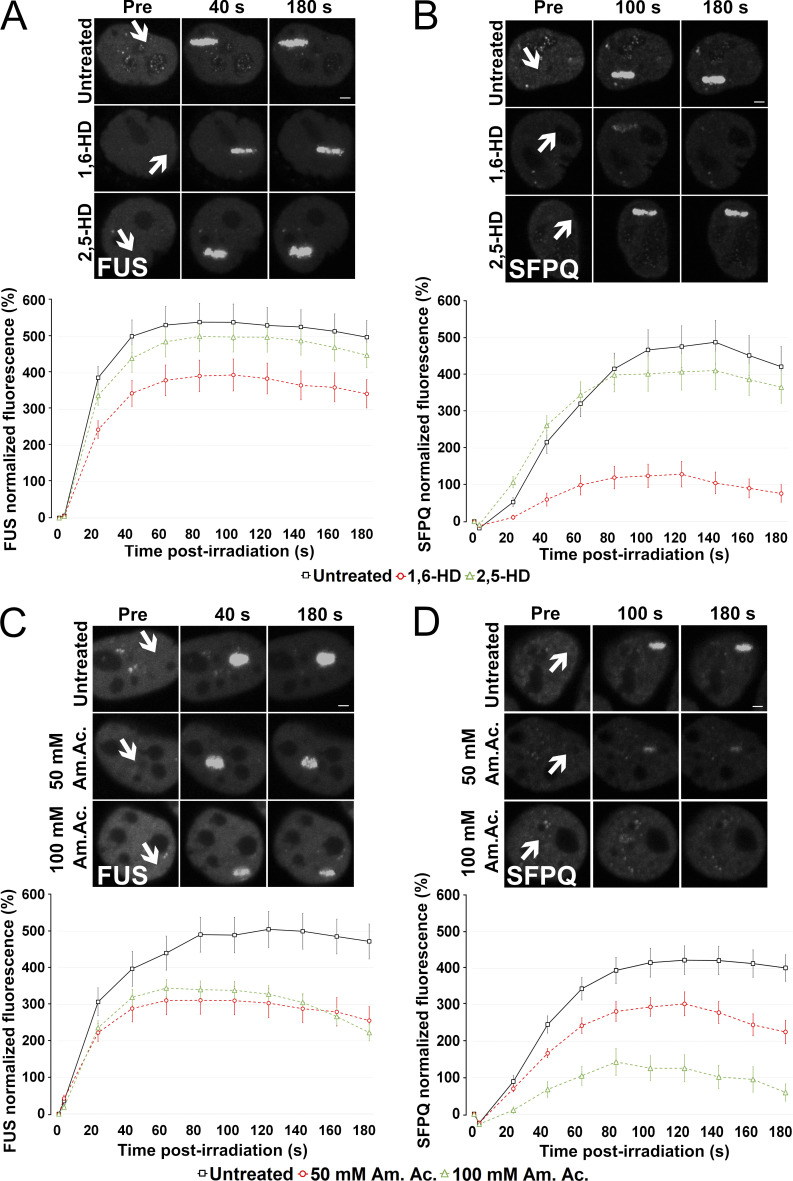
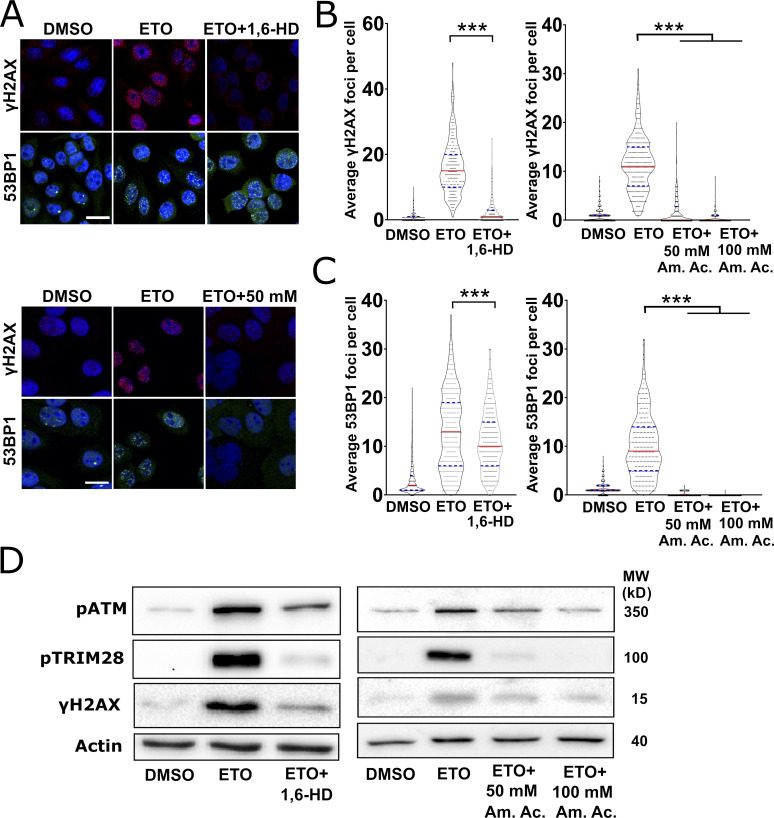
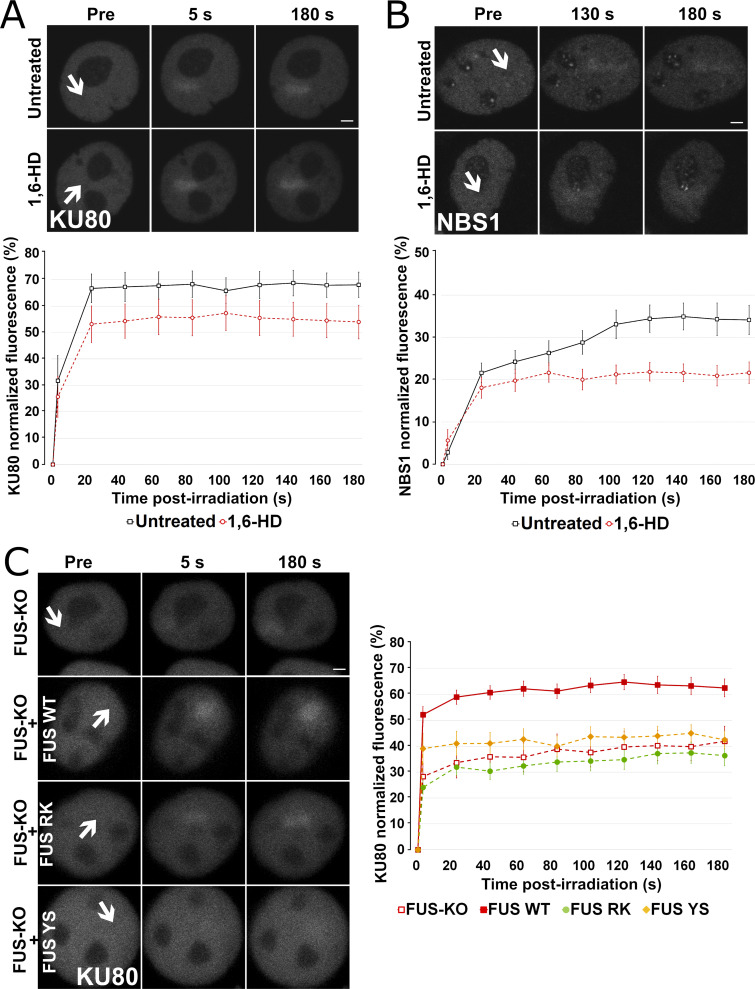
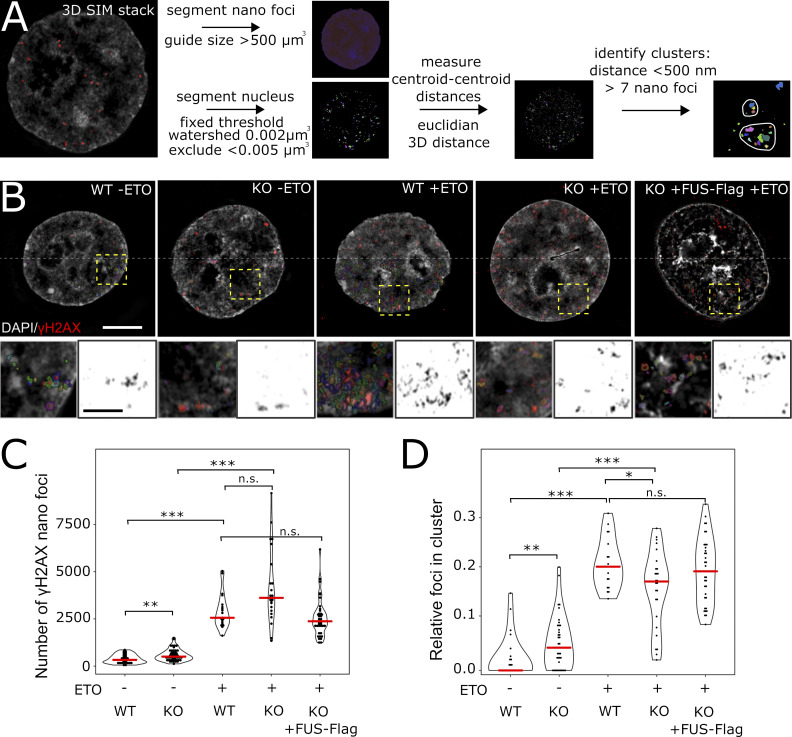
RNA-binding proteins (RBPs) are emerging as important effectors of the cellular DNA damage response (DDR). The RBP FUS is implicated in RNA metabolism and DNA repair, and it undergoes reversible liquid-liquid phase separation (LLPS) in vitro. Here, we demonstrate that FUS-dependent LLPS is necessary for the initiation of the DDR. Using laser microirradiation in FUS-knockout cells, we show that FUS is required for the recruitment to DNA damage sites of the DDR factors KU80, NBS1, and 53BP1 and of SFPQ, another RBP implicated in the DDR. The relocation of KU80, NBS1, and SFPQ is similarly impaired by LLPS inhibitors, or LLPS-deficient FUS variants. We also show that LLPS is necessary for efficient γH2AX foci formation. Finally, using superresolution structured illumination microscopy, we demonstrate that the absence of FUS impairs the proper arrangement of γH2AX nanofoci into higher-order clusters. These findings demonstrate the early requirement for FUS-dependent LLPS in the activation of the DDR and the proper assembly of DSB repair complexes.
© 2021 Levone et al.
Figures





References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
