

Local accumulation of extracellular matrix regulates global morphogenetic patterning in the developing mammary gland
- PMID: 33705716
- PMCID: PMC8119325
- DOI: 10.1016/j.cub.2021.02.015
Local accumulation of extracellular matrix regulates global morphogenetic patterning in the developing mammary gland
Abstract
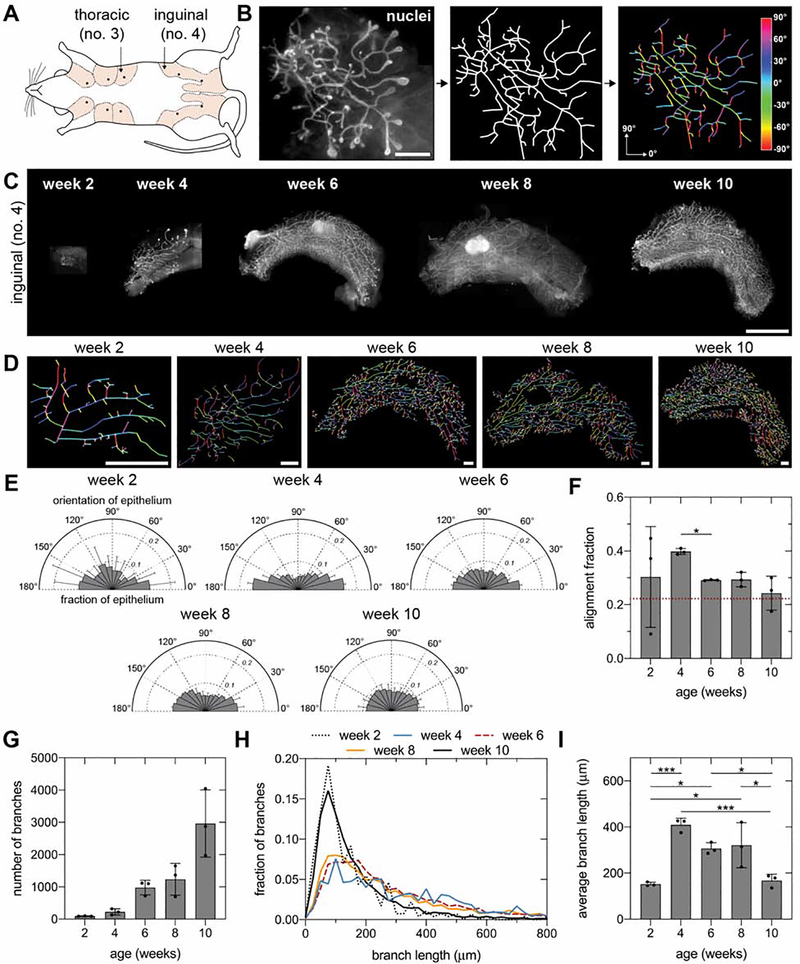
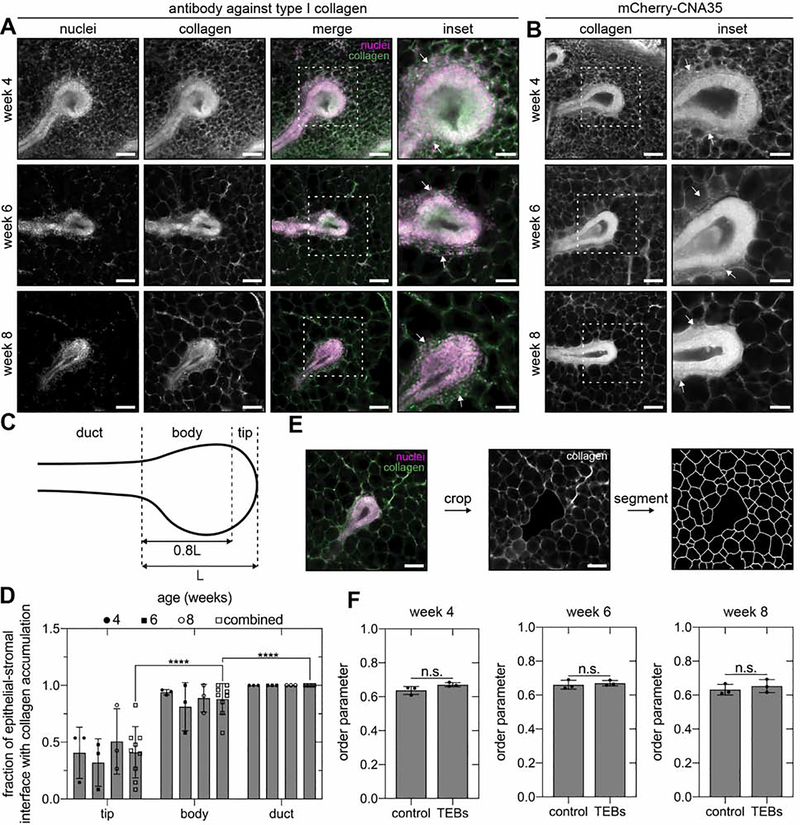
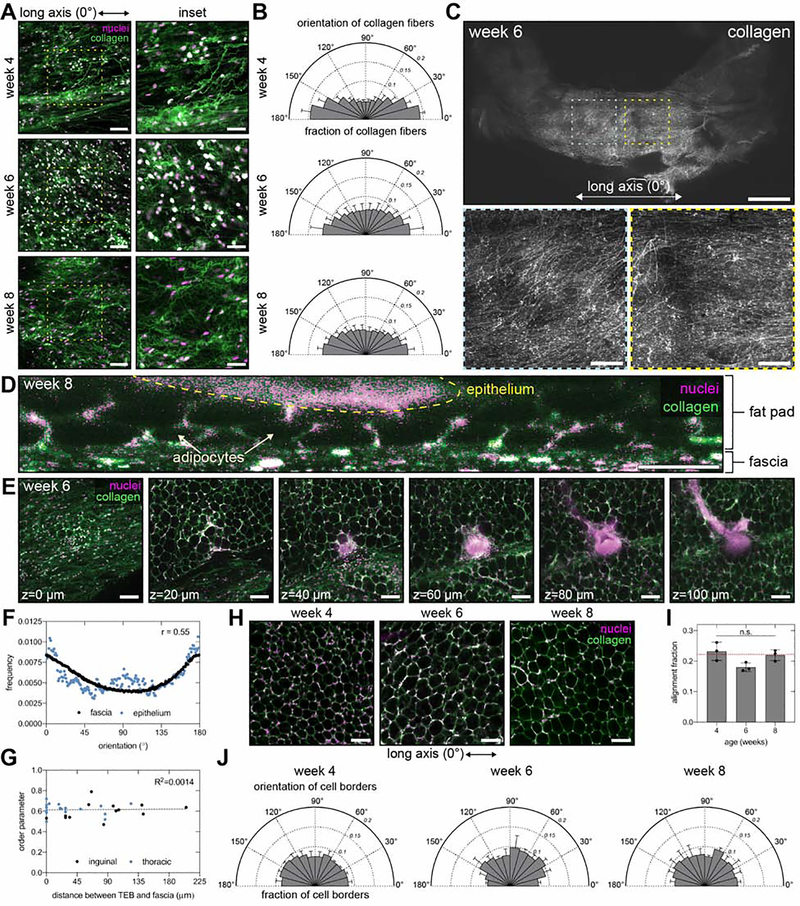
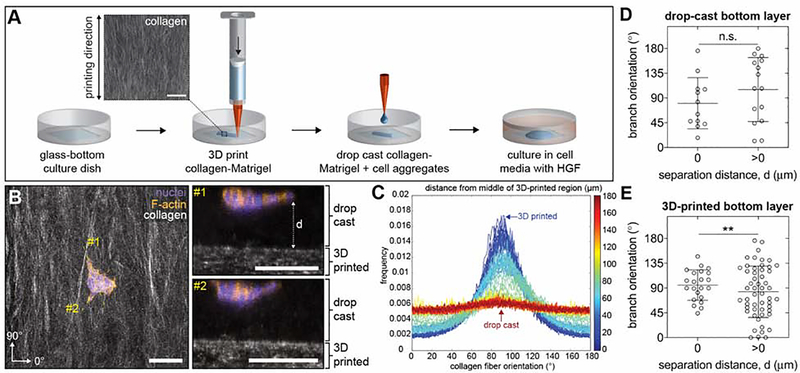
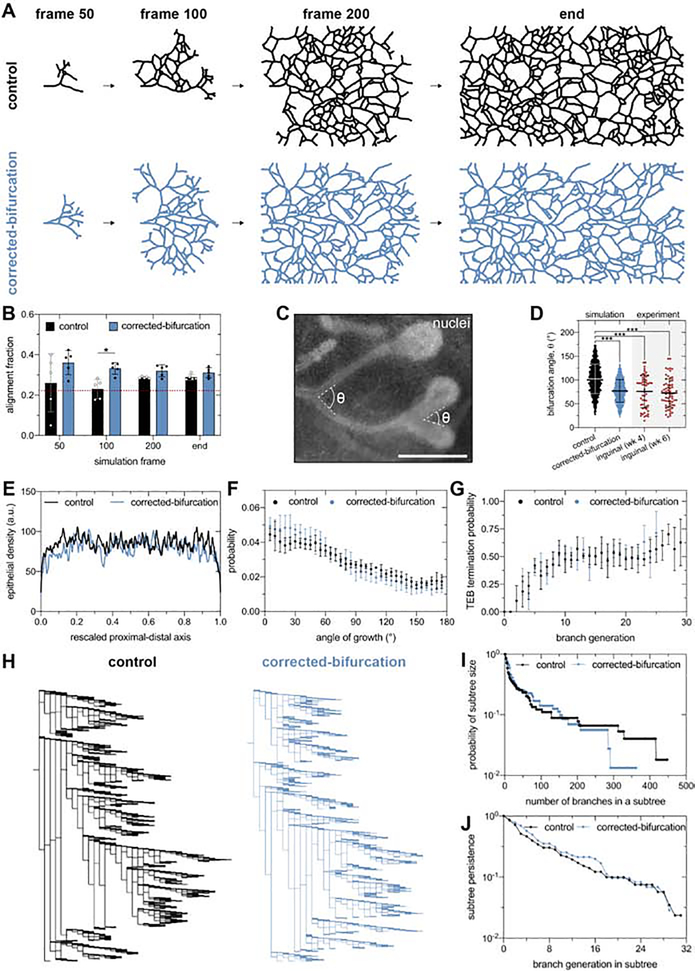
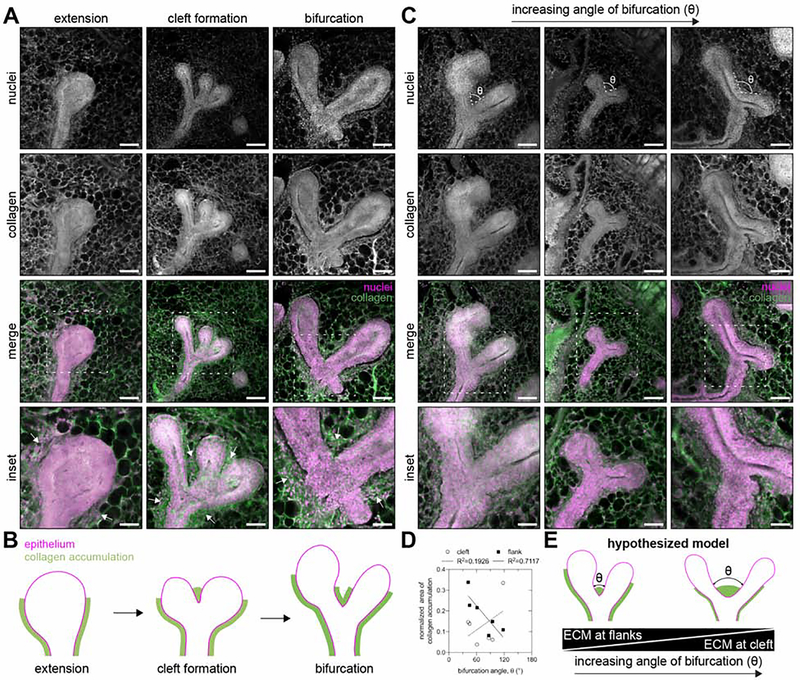
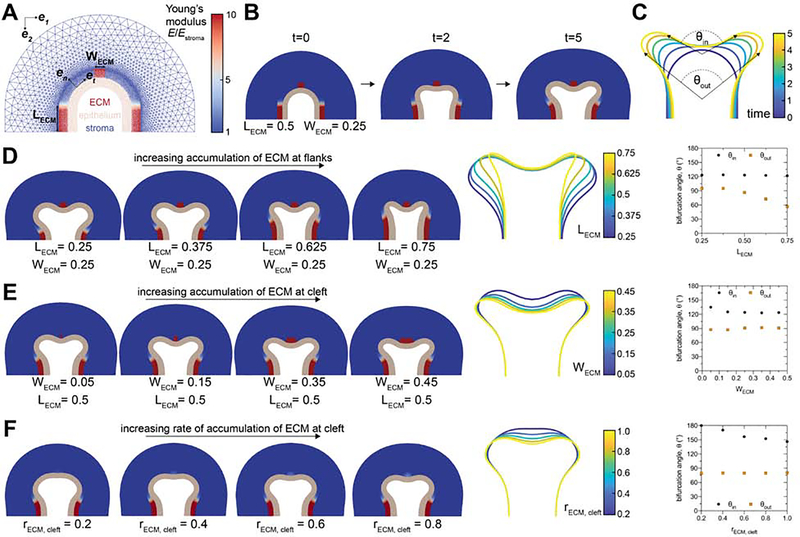
The tree-like pattern of the mammary epithelium is formed during puberty through a process known as branching morphogenesis. Although mammary epithelial branching is stochastic and generates an epithelial tree with a random pattern of branches, the global orientation of the developing epithelium is predictably biased along the long axis of the gland. Here, we combine analysis of pubertal mouse mammary glands, a three-dimensional (3D)-printed engineered tissue model, and computational models of morphogenesis to investigate the origin and the dynamics of the global bias in epithelial orientation during pubertal mammary development. Confocal microscopy analysis revealed that a global bias emerges in the absence of pre-aligned networks of type I collagen in the fat pad and is maintained throughout pubertal development until the widespread formation of lateral branches. Using branching and annihilating random walk simulations, we found that the angle of bifurcation of terminal end buds (TEBs) dictates both the dynamics and the extent of the global bias in epithelial orientation. Our experimental and computational data demonstrate that a local increase in stiffness from the accumulation of extracellular matrix, which constrains the angle of bifurcation of TEBs, is sufficient to pattern the global orientation of the developing mammary epithelium. These data reveal that local mechanical properties regulate the global pattern of mammary epithelial branching and may provide new insight into the global patterning of other branched epithelia.
Keywords: biofabrication; collective migration; finite element method; mechanical stress; mechanotransduction; morphodynamics; tissue engineering.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Daniel CW, Robinson S, and Silberstein GB (1996). The role of TGF-β in patterning and growth of the mammary ductal tree. J. Mammary Gland Biol. Neoplasia 1, 331–341. - PubMed
-
- Gjorevski N, and Nelson CM (2011). Integrated morphodynamic signalling of the mammary gland. Nat. Rev. Mol. Cell Biol 12, 581–593. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
