

Efficient bioconversion of raspberry ketone in Escherichia coli using fatty acids feedstocks
- PMID: 33706766
- PMCID: PMC7953670
- DOI: 10.1186/s12934-021-01551-0
Efficient bioconversion of raspberry ketone in Escherichia coli using fatty acids feedstocks
Abstract
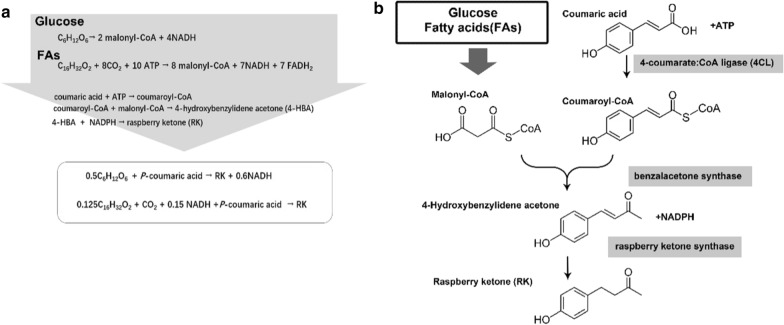
Background: Phenylpropanoid including raspberry ketone, is a kind of important natural plant product and widely used in pharmaceuticals, chemicals, cosmetics, and healthcare products. Bioproduction of phenylpropanoid in Escherichia coli and other microbial cell factories is an attractive approach considering the low phenylpropanoid contents in plants. However, it is usually difficult to produce high titer phenylpropanoid production when fermentation using glucose as carbon source. Developing novel bioprocess using alternative sources might provide a solution to this problem. In this study, typical phenylpropanoid raspberry ketone was used as the target product to develop a biosynthesis pathway for phenylpropanoid production from fatty acids, a promising alternative low-cost feedstock.
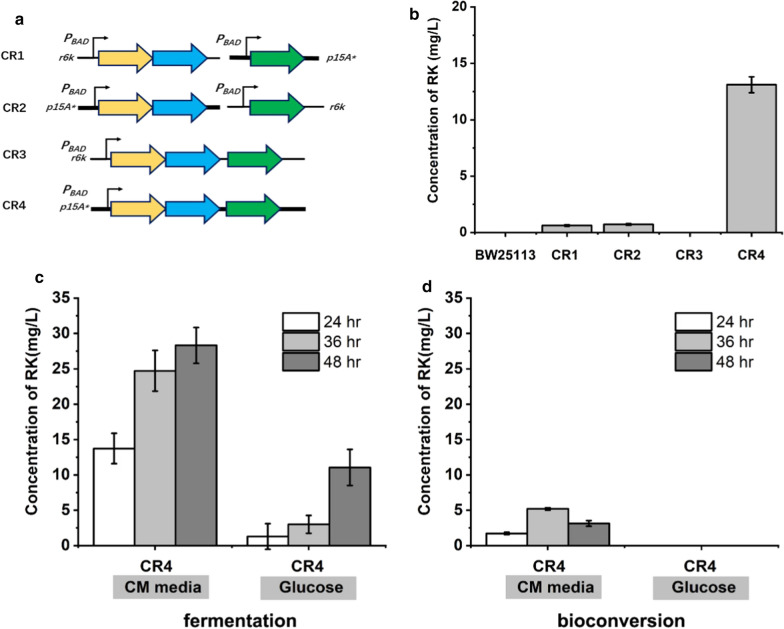
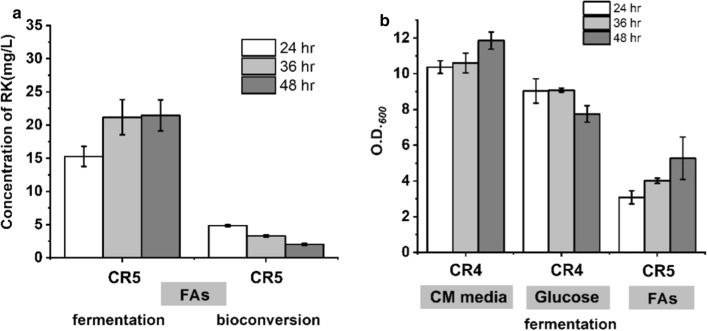
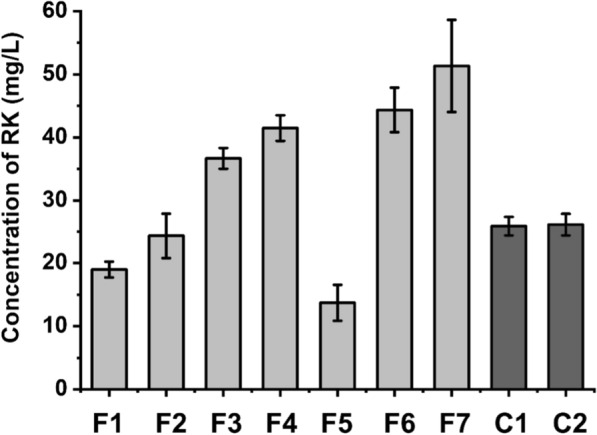
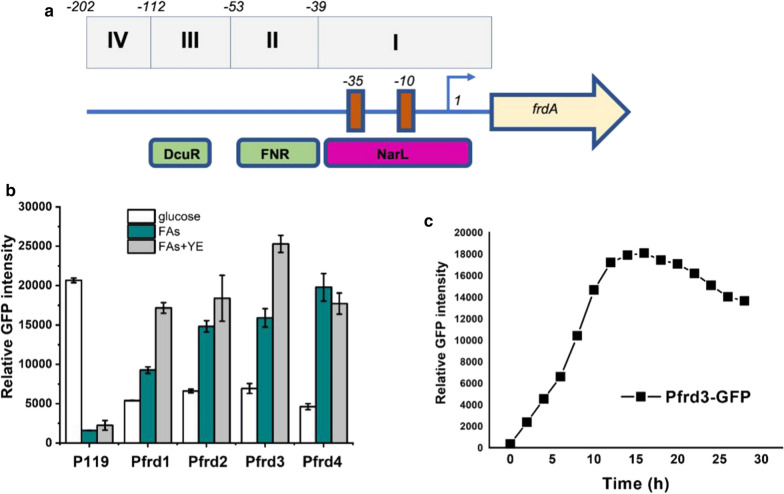
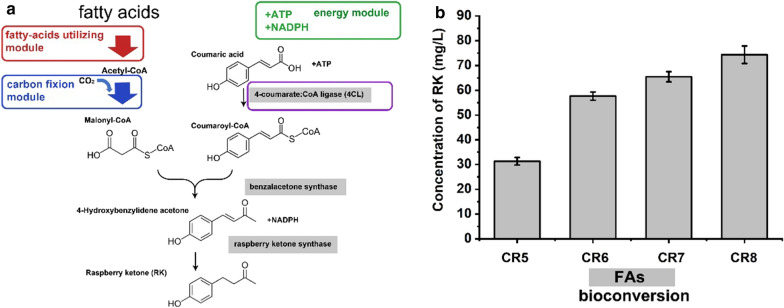
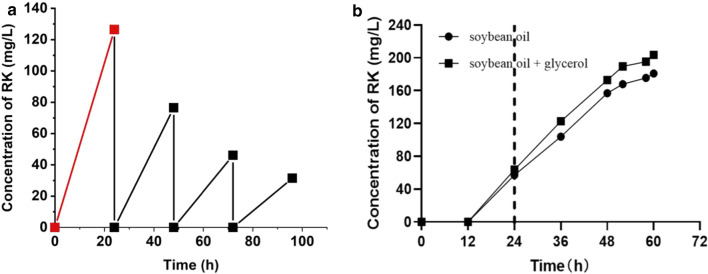
Results: A raspberry ketone biosynthesis module was developed and optimized by introducing 4-coumarate-CoA ligase (4CL), benzalacetone synthase (BAS), and raspberry ketone reductase (RZS) in Escherichia coli strains CR1-CR4. Then strain CR5 was developed by introducing raspberry ketone biosynthesis module into a fatty acids-utilization chassis FA09 to achieve production of raspberry ketone from fatty acids feedstock. However, the production of raspberry ketone was still limited by the low biomass and unable to substantiate whole-cell bioconversion process. Thus, a process by coordinately using fatty-acids and glycerol was developed. In addition, we systematically screened and optimized fatty acids-response promoters. The optimized promoter Pfrd3 was then successfully used for the efficient expression of key enzymes of raspberry ketone biosynthesis module during bioconversion from fatty acids. The final engineered strain CR8 could efficiently produce raspberry ketone repeatedly using bioconversion from fatty acids feedstock strategy, and was able to produce raspberry ketone to a concentration of 180.94 mg/L from soybean oil in a 1-L fermentation process.
Conclusion: Metabolically engineered Escherichia coli strains were successfully developed for raspberry ketone production from fatty acids using several strategies, including optimization of bioconversion process and fine-tuning key enzyme expression. This study provides an essential reference to establish the low-cost biological manufacture of phenylpropanoids compounds.
Keywords: Bioconversion; Escherichia coli; Fatty acids feedstock; Phenylpropanoids; Raspberry ketone.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
