

The transcription factor Bcl11b promotes both canonical and adaptive NK cell differentiation
- PMID: 33712472
- PMCID: PMC8274449
- DOI: 10.1126/sciimmunol.abc9801
The transcription factor Bcl11b promotes both canonical and adaptive NK cell differentiation
Abstract
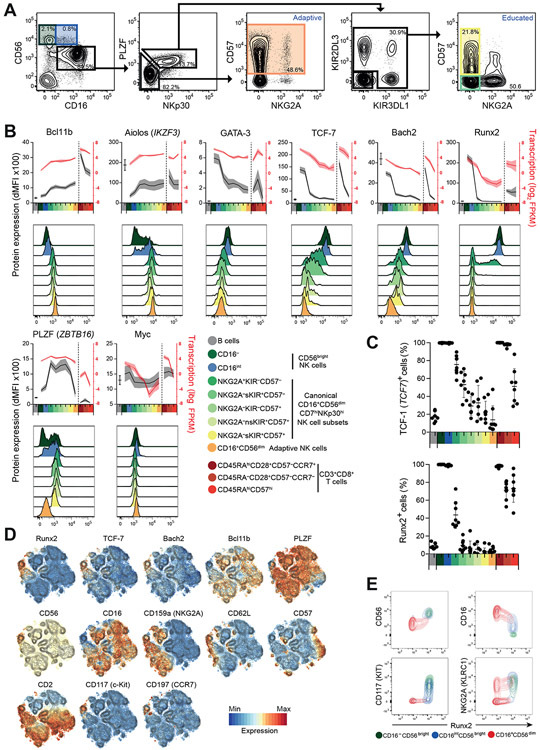
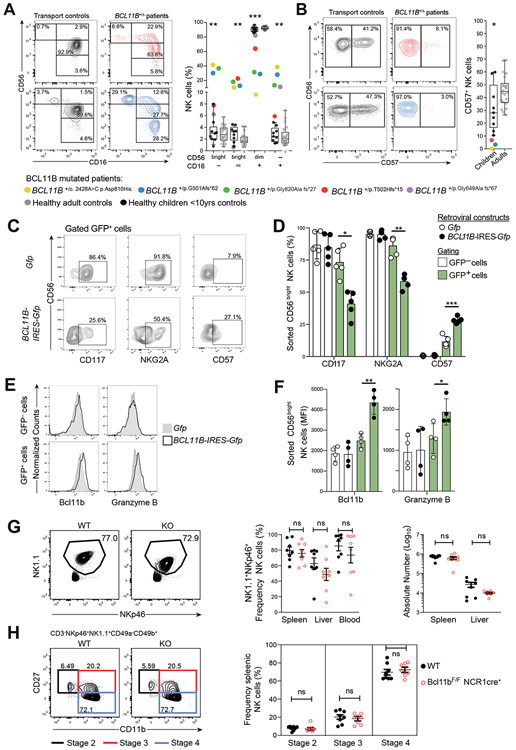
Epigenetic landscapes can provide insight into regulation of gene expression and cellular diversity. Here, we examined the transcriptional and epigenetic profiles of seven human blood natural killer (NK) cell populations, including adaptive NK cells. The BCL11B gene, encoding a transcription factor (TF) essential for T cell development and function, was the most extensively regulated, with expression increasing throughout NK cell differentiation. Several Bcl11b-regulated genes associated with T cell signaling were specifically expressed in adaptive NK cell subsets. Regulatory networks revealed reciprocal regulation at distinct stages of NK cell differentiation, with Bcl11b repressing RUNX2 and ZBTB16 in canonical and adaptive NK cells, respectively. A critical role for Bcl11b in driving NK cell differentiation was corroborated in BCL11B-mutated patients and by ectopic Bcl11b expression. Moreover, Bcl11b was required for adaptive NK cell responses in a murine cytomegalovirus model, supporting expansion of these cells. Together, we define the TF regulatory circuitry of human NK cells and uncover a critical role for Bcl11b in promoting NK cell differentiation and function.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures






Comment in
-
A natural killer's hike through epigenetic landscapes.Sci Immunol. 2021 Mar 12;6(57):eabf8019. doi: 10.1126/sciimmunol.abf8019. Sci Immunol. 2021. PMID: 33712474
References
-
- Spits H, Artis D, Colonna M, Diefenbach A, Di Santo JP, Eberl G, Koyasu S, Locksley RM, McKenzie AN, Mebius RE, Powrie F, Vivier E, Innate lymphoid cells--a proposal for uniform nomenclature. Nature reviews. Immunology 13, 145–149 (2013). - PubMed
-
- Cannon MJ, Congenital cytomegalovirus (CMV) epidemiology and awareness. J Clin Virol 46 Suppl 4, S6–10 (2009). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
