

Deep-sea microbes as tools to refine the rules of innate immune pattern recognition
- PMID: 33712473
- PMCID: PMC8367048
- DOI: 10.1126/sciimmunol.abe0531
Deep-sea microbes as tools to refine the rules of innate immune pattern recognition
Abstract
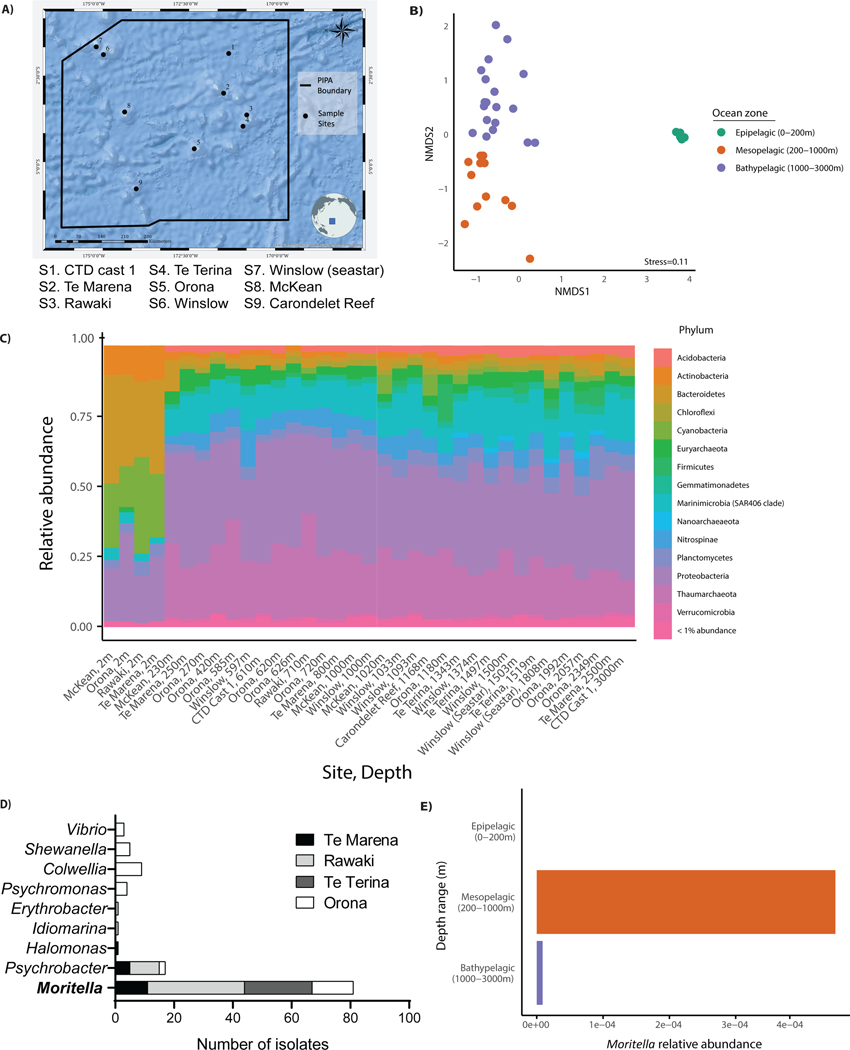
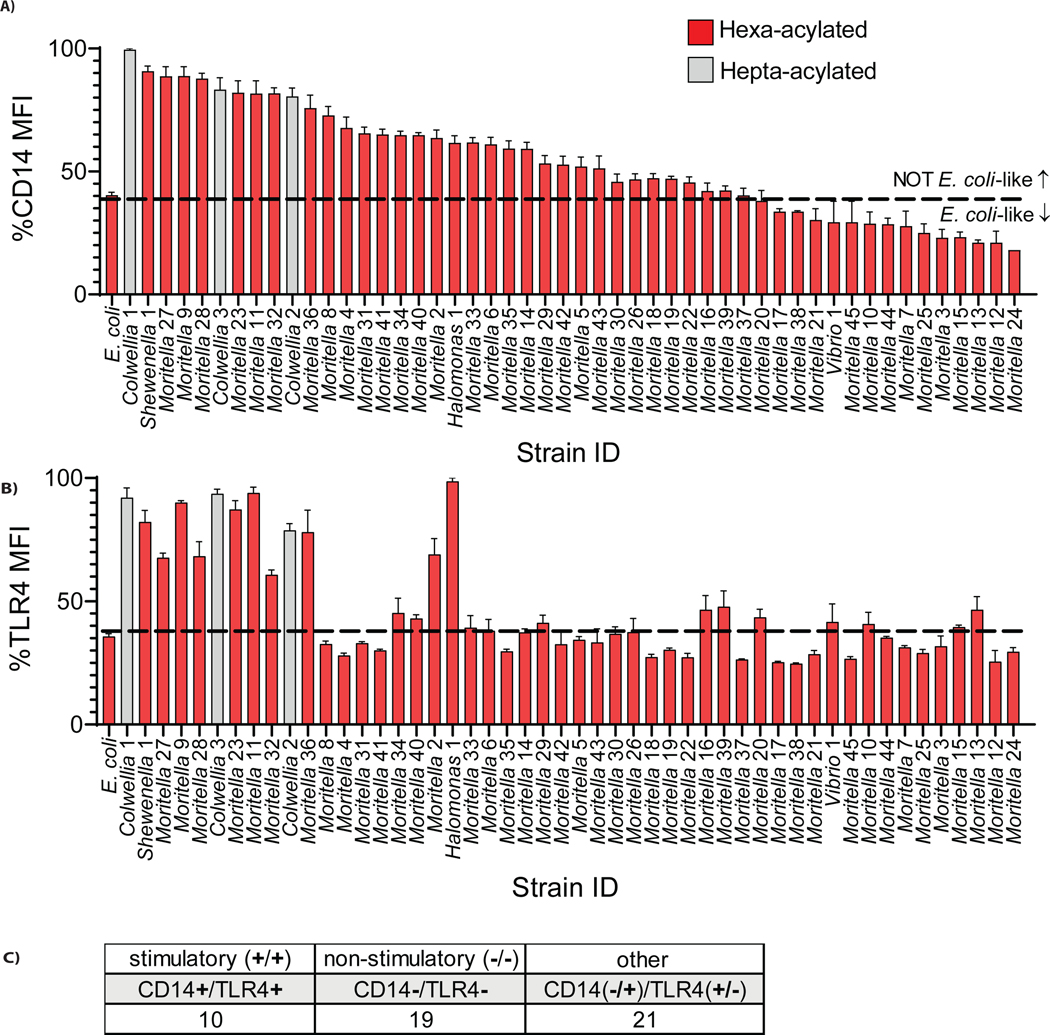
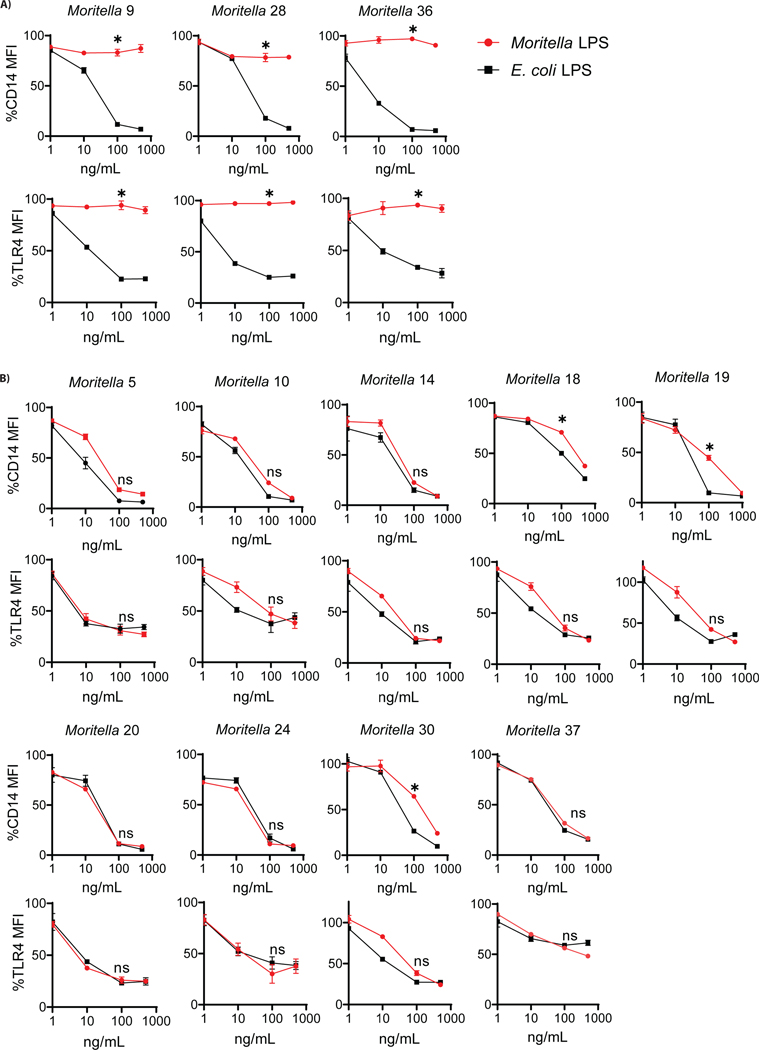
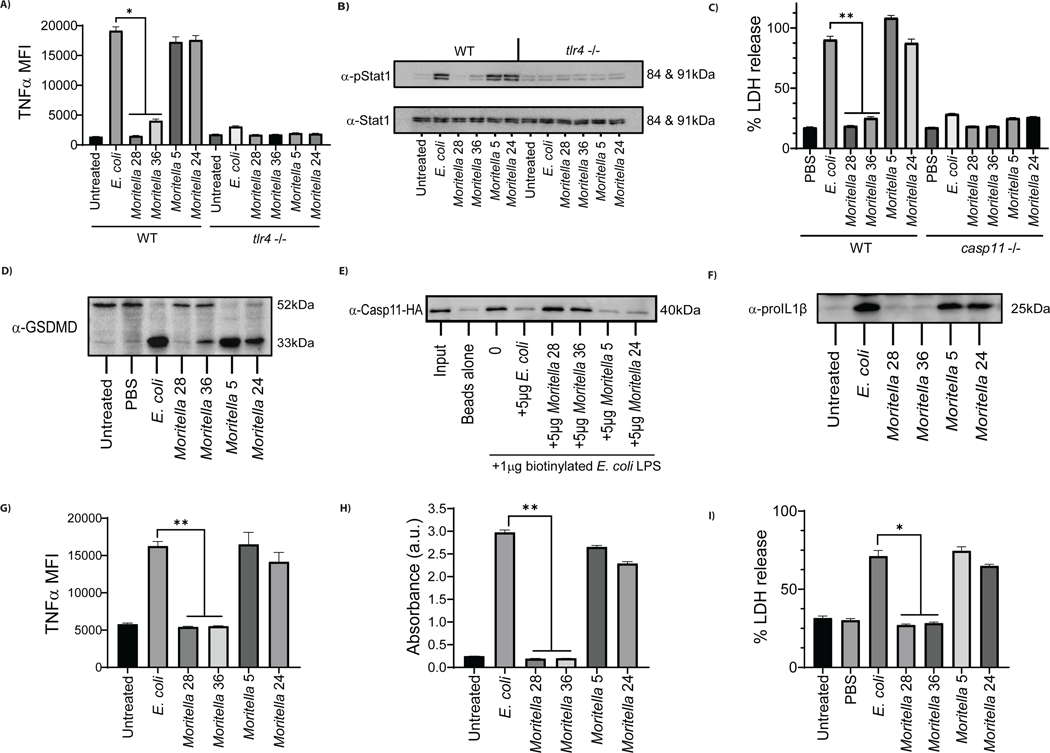
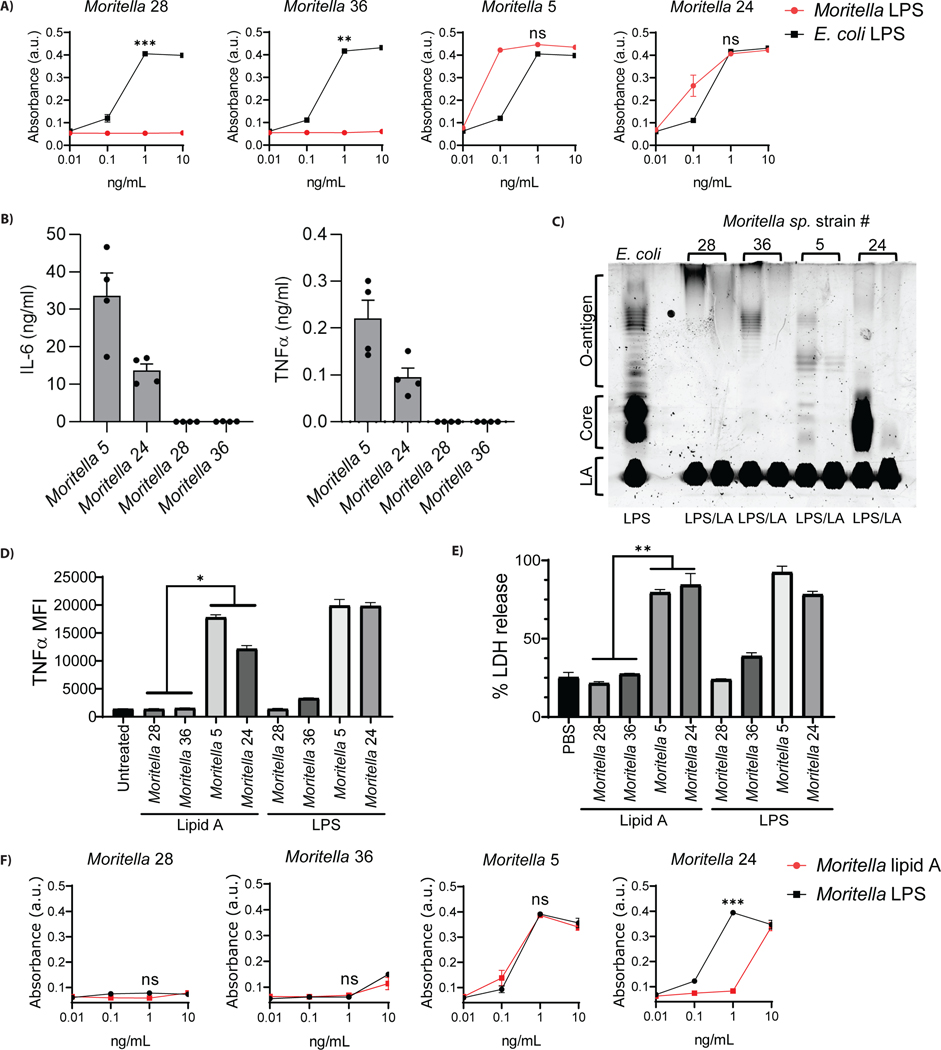
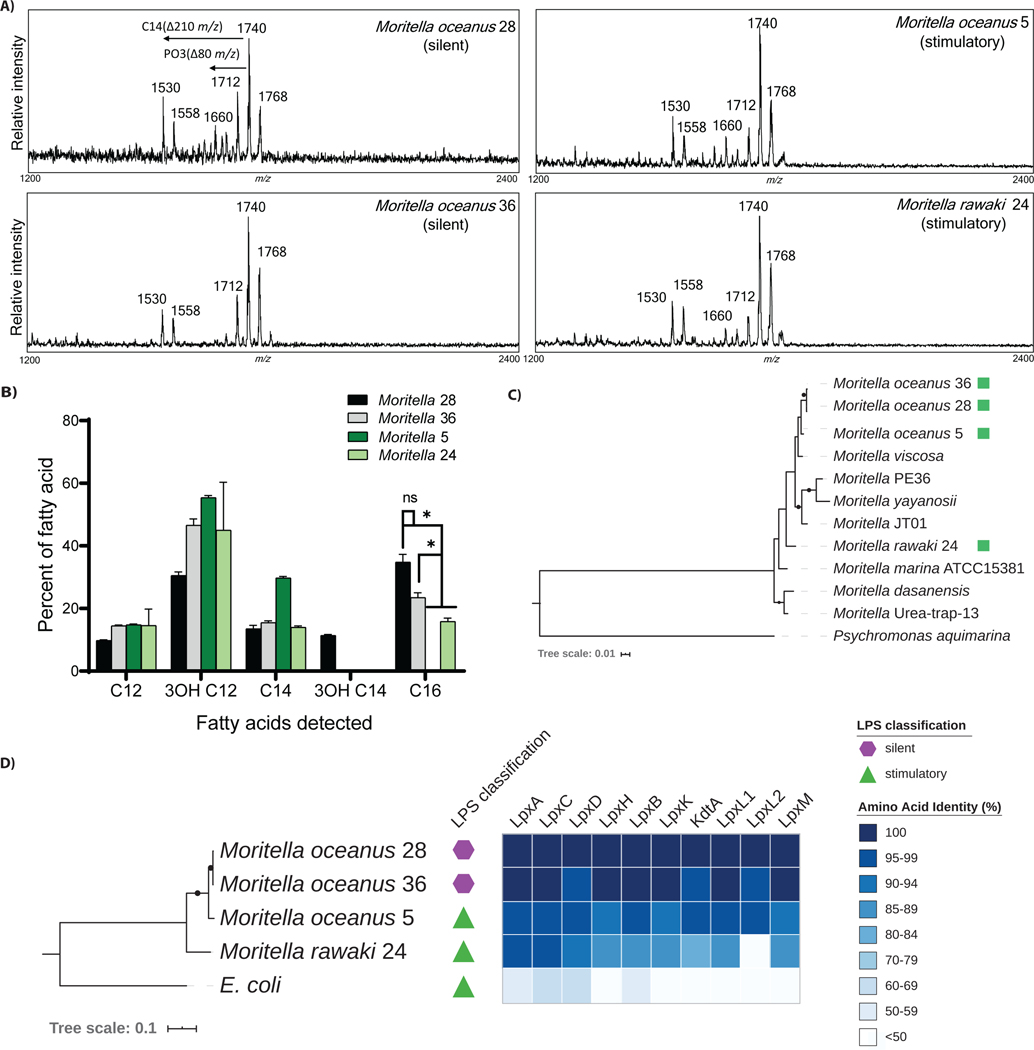
The assumption of near-universal bacterial detection by pattern recognition receptors is a foundation of immunology. The limits of this pattern recognition concept, however, remain undefined. As a test of this hypothesis, we determined whether mammalian cells can recognize bacteria that they have never had the natural opportunity to encounter. These bacteria were cultivated from the deep Pacific Ocean, where the genus Moritella was identified as a common constituent of the culturable microbiota. Most deep-sea bacteria contained cell wall lipopolysaccharide (LPS) structures that were expected to be immunostimulatory, and some deep-sea bacteria activated inflammatory responses from mammalian LPS receptors. However, LPS receptors were unable to detect 80% of deep-sea bacteria examined, with LPS acyl chain length being identified as a potential determinant of immunosilence. The inability of immune receptors to detect most bacteria from a different ecosystem suggests that pattern recognition strategies may be defined locally, not globally.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Competing interests
Boston University and Boston Children’s Hospital have filed a patent application entitled “Immunomodulatory lipopolysaccharide compositions” with R.D.R, A.E.G., A.T. and J.C.K as inventors. J.C.K. holds equity and consults for IFM Therapeutics, Quench Bio and Corner Therapeutics. None of these relationships influenced the work performed in this study. The other authors declare that they have no competing interests.
Figures
References
-
- Janeway CA Jr., Approaching the asymptote? Evolution and revolution in immunology. Cold Spring Harbor symposia on quantitative biology 54 Pt 1, 1–13 (1989). - PubMed
-
- Medzhitov R, Janeway CA, Innate immunity: impact on the adaptive immune response. Current Opinion in Immunology 9, 4–9 (1997). - PubMed
-
- C. A. Janeway, Jr., R. Medzhitov, Innate immune recognition. Annual review of immunology 20, 197–216 (2002). - PubMed
-
- Steimle A, Autenrieth IB, Frick J-S, Structure and function: Lipid A modifications in commensals and pathogens. International Journal of Medical Microbiology 306, 290–301 (2016). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
