

ADAR1 RNA editing enzyme regulates R-loop formation and genome stability at telomeres in cancer cells
- PMID: 33712600
- PMCID: PMC7955049
- DOI: 10.1038/s41467-021-21921-x
ADAR1 RNA editing enzyme regulates R-loop formation and genome stability at telomeres in cancer cells
Abstract
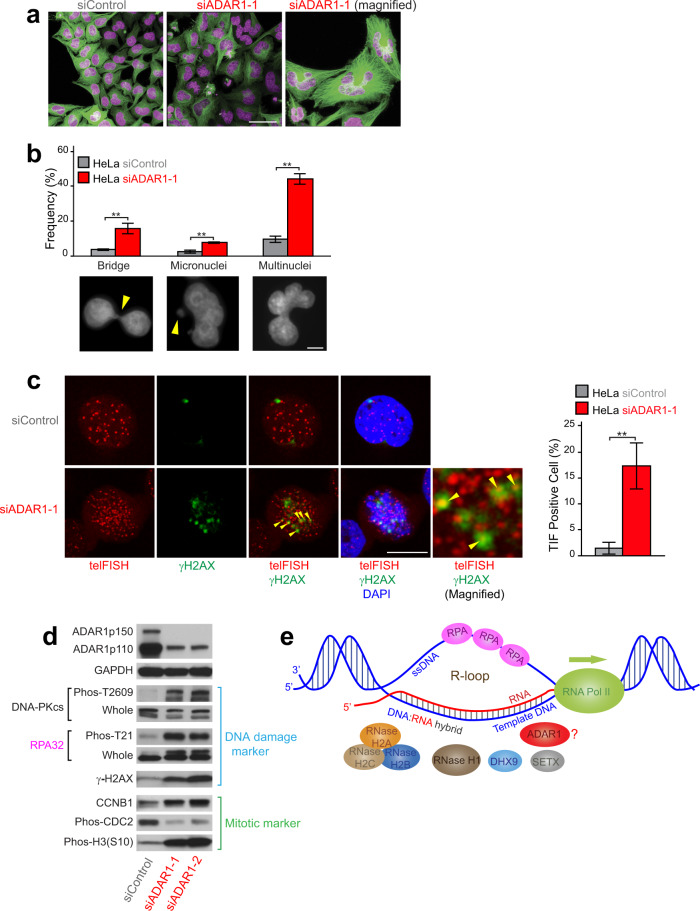
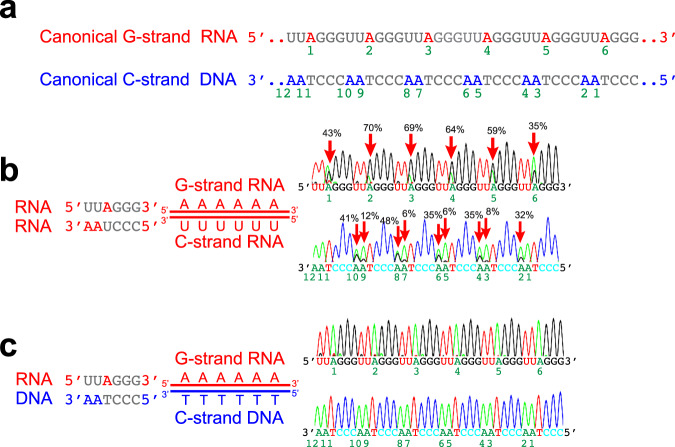
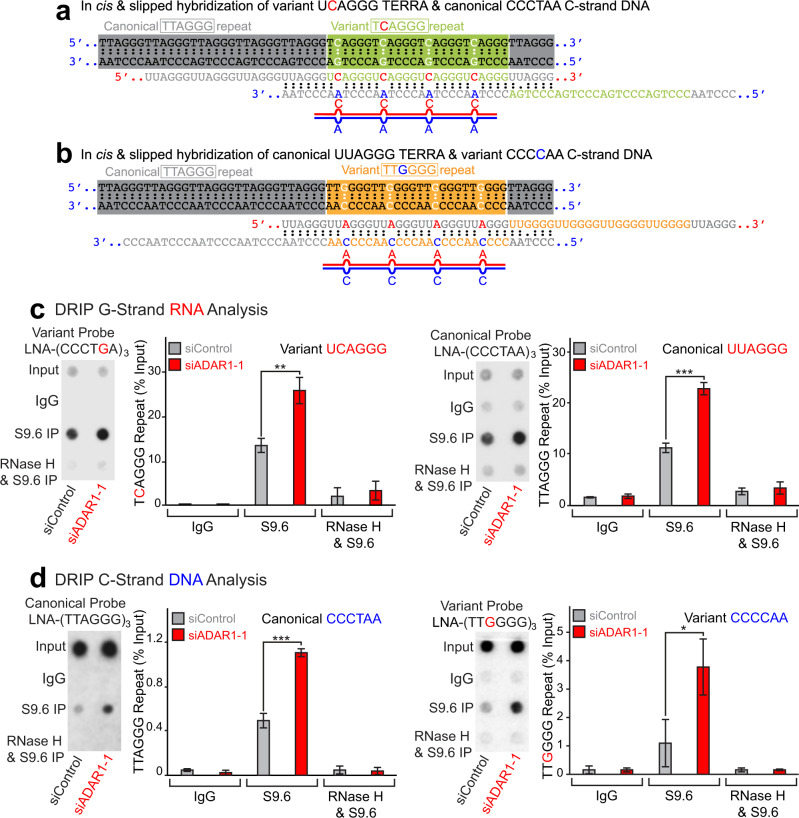
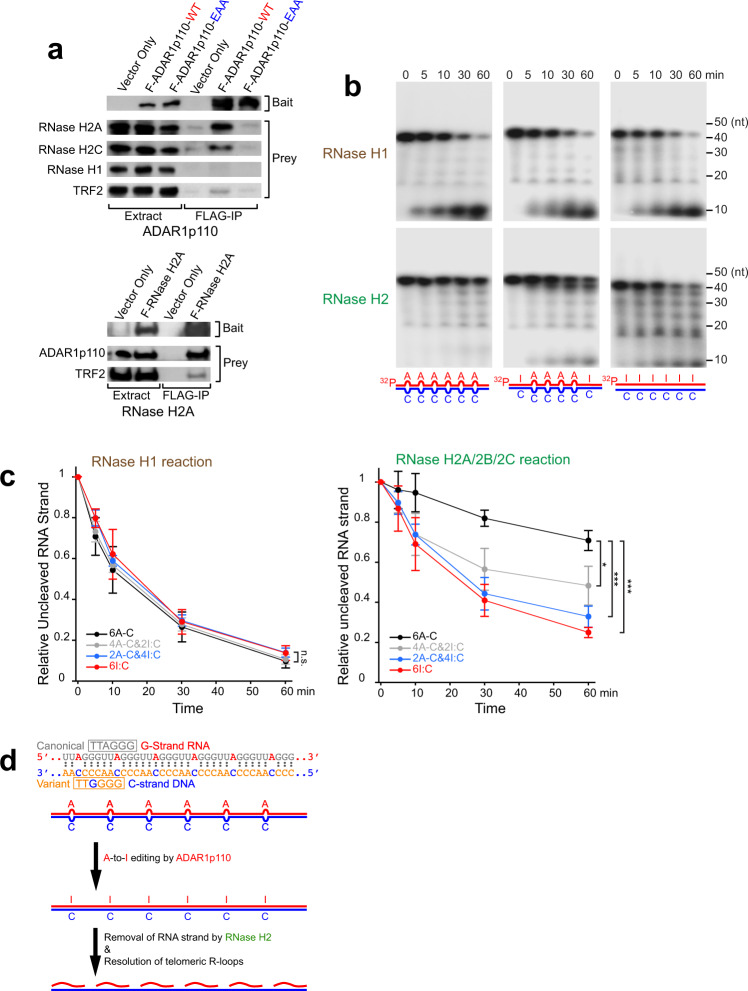
ADAR1 is involved in adenosine-to-inosine RNA editing. The cytoplasmic ADAR1p150 edits 3'UTR double-stranded RNAs and thereby suppresses induction of interferons. Loss of this ADAR1p150 function underlies the embryonic lethality of Adar1 null mice, pathogenesis of the severe autoimmune disease Aicardi-Goutières syndrome, and the resistance developed in cancers to immune checkpoint blockade. In contrast, the biological functions of the nuclear-localized ADAR1p110 remain largely unknown. Here, we report that ADAR1p110 regulates R-loop formation and genome stability at telomeres in cancer cells carrying non-canonical variants of telomeric repeats. ADAR1p110 edits the A-C mismatches within RNA:DNA hybrids formed between canonical and non-canonical variant repeats. Editing of A-C mismatches to I:C matched pairs facilitates resolution of telomeric R-loops by RNase H2. This ADAR1p110-dependent control of telomeric R-loops is required for continued proliferation of telomerase-reactivated cancer cells, revealing the pro-oncogenic nature of ADAR1p110 and identifying ADAR1 as a promising therapeutic target of telomerase positive cancers.
Conflict of interest statement
The authors declare no competing interests.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
