

Identification of eight QTL controlling multiple yield components in a German multi-parental wheat population, including Rht24, WAPO-A1, WAPO-B1 and genetic loci on chromosomes 5A and 6A
- PMID: 33712876
- PMCID: PMC8081691
- DOI: 10.1007/s00122-021-03781-7
Identification of eight QTL controlling multiple yield components in a German multi-parental wheat population, including Rht24, WAPO-A1, WAPO-B1 and genetic loci on chromosomes 5A and 6A
Abstract
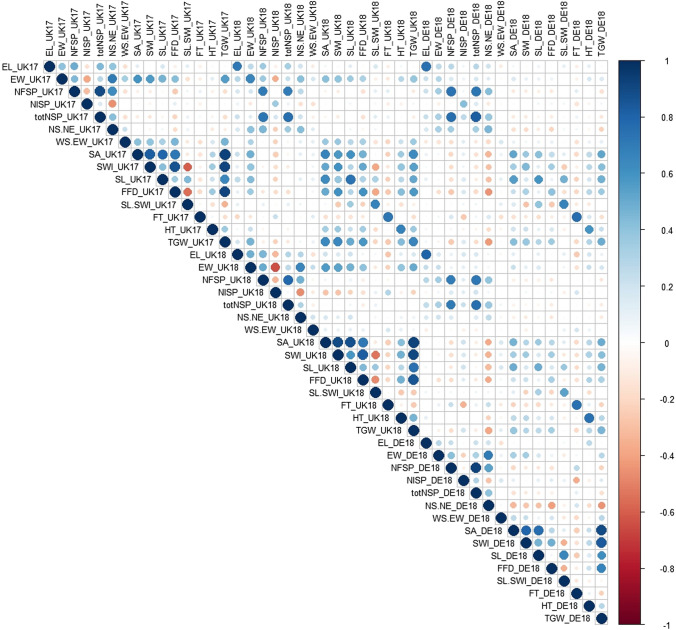
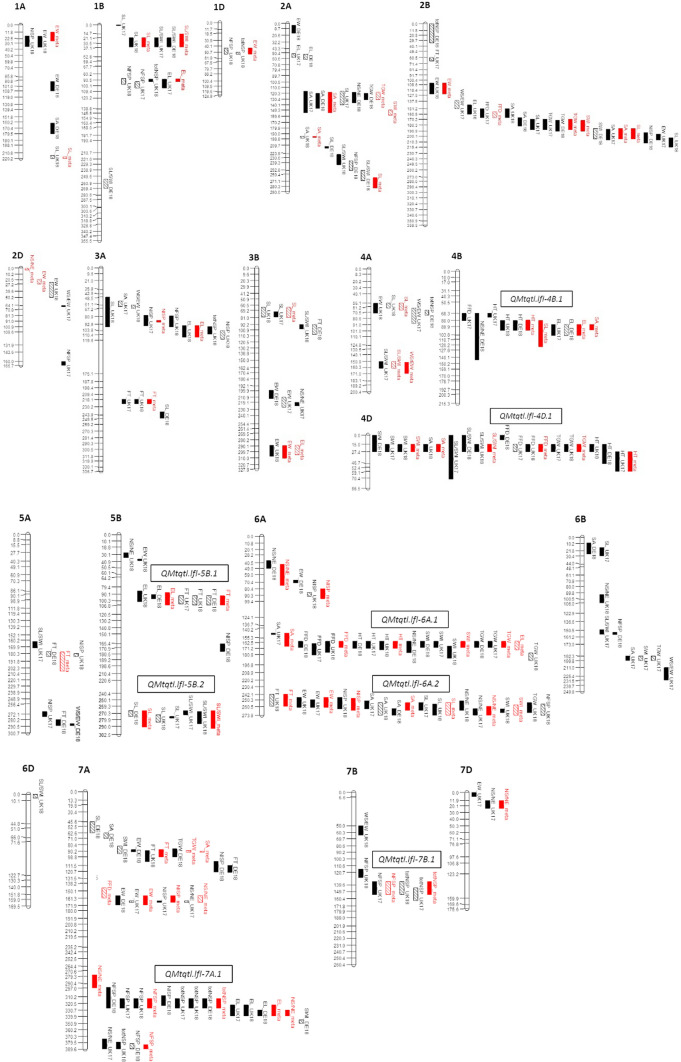
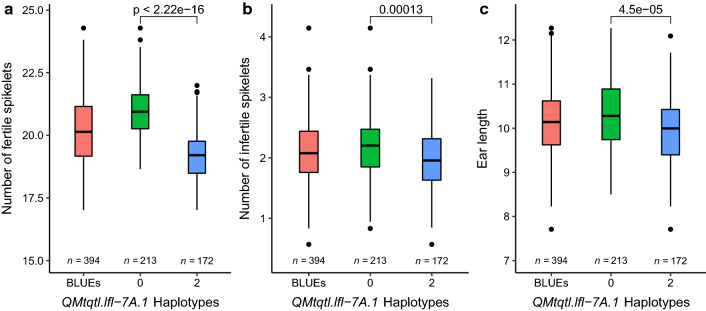
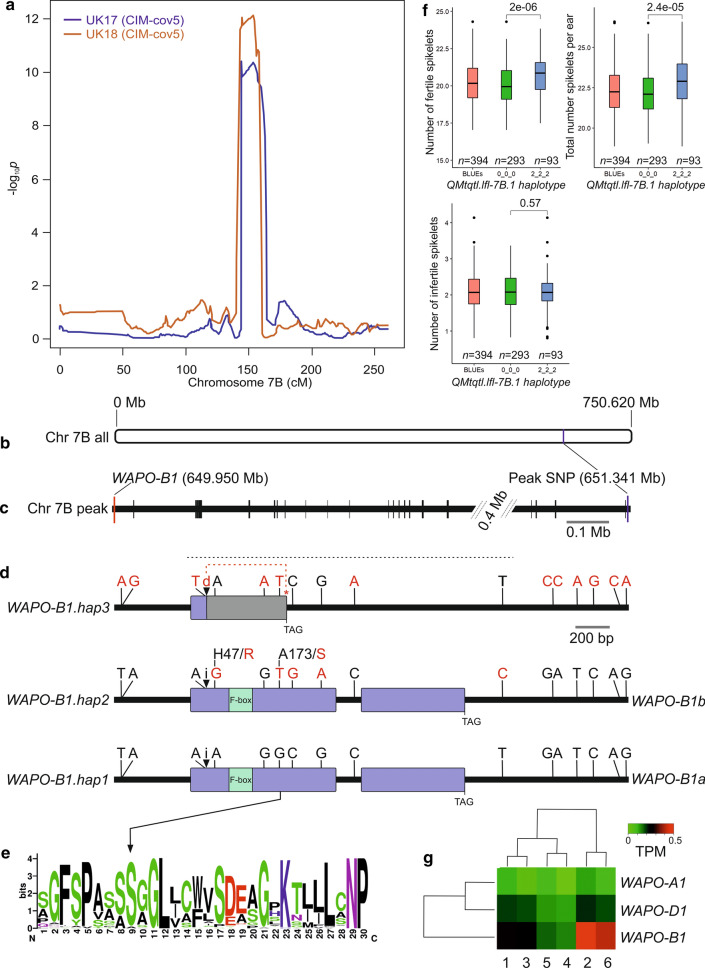
Quantitative trait locus (QTL) mapping of 15 yield component traits in a German multi-founder population identified eight QTL each controlling ≥2 phenotypes, including the genetic loci Rht24, WAPO-A1 and WAPO-B1. Grain yield in wheat (Triticum aestivum L.) is a polygenic trait representing the culmination of many developmental processes and their interactions with the environment. Toward maintaining genetic gains in yield potential, 'reductionist approaches' are commonly undertaken by which the genetic control of yield components, that collectively determine yield, are established. Here we use an eight-founder German multi-parental wheat population to investigate the genetic control and phenotypic trade-offs between 15 yield components. Increased grains per ear was significantly positively correlated with the number of fertile spikelets per ear and negatively correlated with the number of infertile spikelets. However, as increased grain number and fertile spikelet number per ear were significantly negatively correlated with thousand grain weight, sink strength limitations were evident. Genetic mapping identified 34 replicated quantitative trait loci (QTL) at two or more test environments, of which 24 resolved into eight loci each controlling two or more traits-termed here 'multi-trait QTL' (MT-QTL). These included MT-QTL associated with previously cloned genes controlling semi-dwarf plant stature, and with the genetic locus Reduced height 24 (Rht24) that further modulates plant height. Additionally, MT-QTL controlling spikelet number traits were located to chromosome 7A encompassing the gene WHEAT ORTHOLOG OF APO1 (WAPO-A1), and to its homoeologous location on chromosome 7B containing WAPO-B1. The genetic loci identified in this study, particularly those that potentially control multiple yield components, provide future opportunities for the targeted investigation of their underlying genes, gene networks and phenotypic trade-offs, in order to underpin further genetic gains in yield.
Conflict of interest statement
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Figures
References
-
- Altschul SF, Gish W, Miller E, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Bates D, Mächler M, Bolker B, Walker S. Fitting linear mixed-effects models using lme4. J Stat Softw. 2015;67:1–48. doi: 10.18637/jss.v067.i01. - DOI
-
- Bonnet OT. The development of the wheat spike. J Agr Res. 1936;53:445–451.
-
- Brinton J, Simmonds J, Minter F, Leverington-Waite M, Snape J, Uauy C. Increased pericarp cell length underlies a major quantitative trait locus for grain weight in hexaploid wheat. New Phytol. 2017;215:1026–1038. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
