

Pericytes Regulate Cerebral Perfusion through VEGFR1 in Ischemic Stroke
- PMID: 33712886
- PMCID: PMC11421746
- DOI: 10.1007/s10571-021-01071-w
Pericytes Regulate Cerebral Perfusion through VEGFR1 in Ischemic Stroke
Abstract
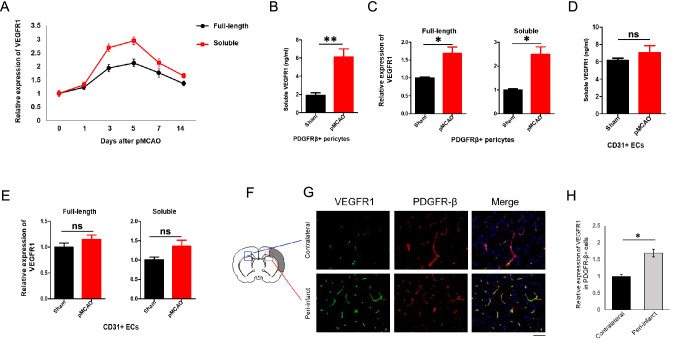
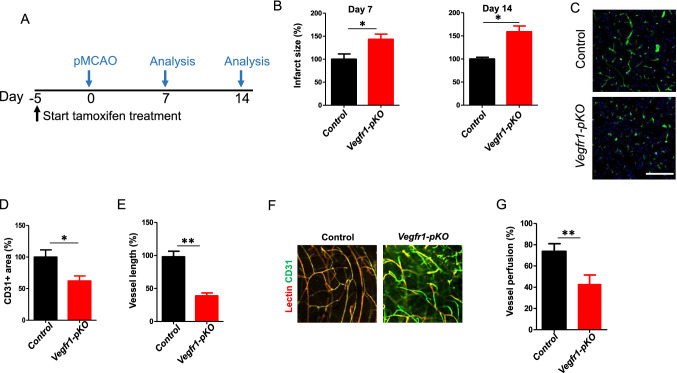
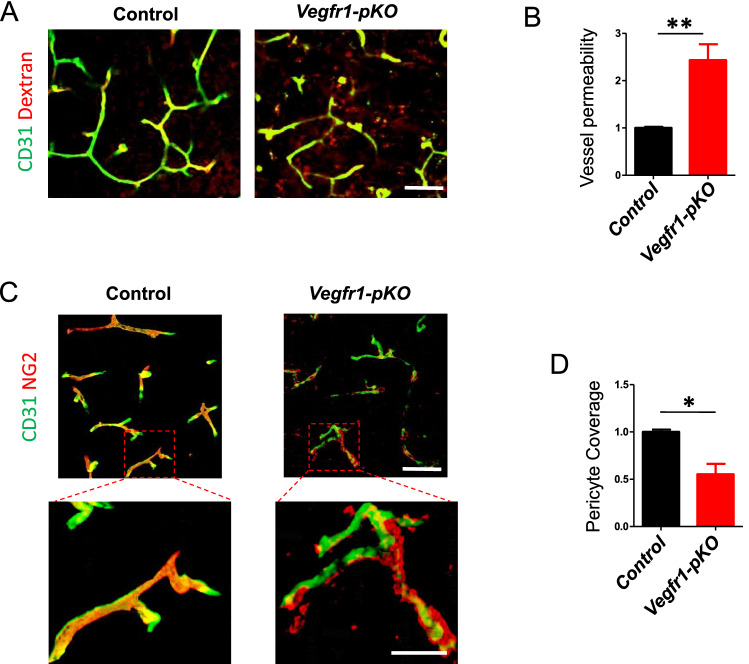
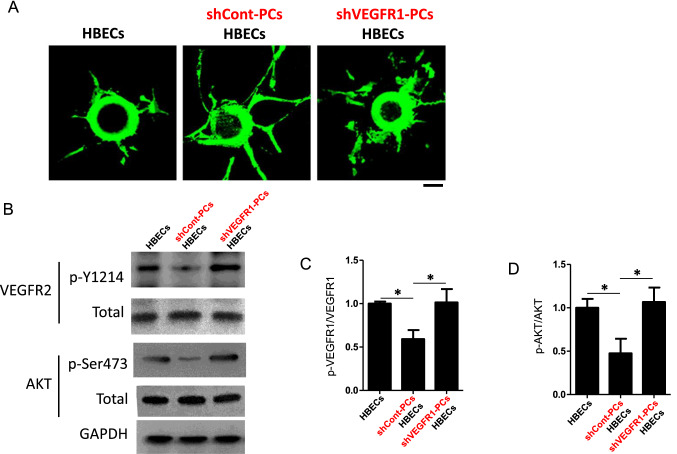
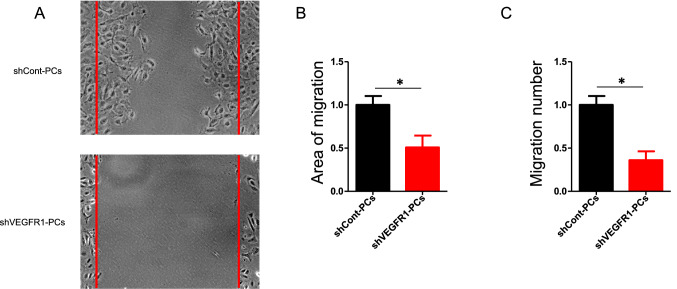
Neurons in the penumbra (the area surrounding ischemic tissue that consists of still viable tissue but with reduced blood flow and oxygen transport) may be rescued following stroke if adequate perfusion is restored in time. It has been speculated that post-stroke angiogenesis in the penumbra can reduce damage caused by ischemia. However, the mechanism for neovasculature formation in the brain remains unclear and vascular-targeted therapies for brain ischemia remain suboptimal. Here, we show that VEGFR1 was highly upregulated in pericytes after stroke. Knockdown of VEGFR1 in pericytes led to increased infarct area and compromised post-ischemia vessel formation. Furthermore, in vitro studies confirmed a critical role for pericyte-derived VEGFR1 in both endothelial tube formation and pericyte migration. Interestingly, our results show that pericyte-derived VEGFR1 has opposite effects on Akt activity in endothelial cells and pericytes. Collectively, these results indicate that pericyte-specific expression of VEGFR1 modulates ischemia-induced vessel formation and vascular integrity in the brain.
Keywords: Cerebrovascular; Ischemia; Pericyte; Stroke; VEGFR1.
© 2021. The Author(s), under exclusive licence to Springer Science+Business Media, LLC, part of Springer Nature.
Conflict of interest statement
The authors have no conflict of interest to declare.
Figures


References
-
- Arenillas JF, Sobrino T, Castillo J, Davalos A (2007) The role of angiogenesis in damage and recovery from ischemic stroke. Curr Treat Options Cardiovasc Med 9(3):205–212 - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
