

Too much of a good thing: Adaption to iron (II) intoxication in Escherichia coli
- PMID: 33717488
- PMCID: PMC7937436
- DOI: 10.1093/emph/eoaa051
Too much of a good thing: Adaption to iron (II) intoxication in Escherichia coli
Abstract
Background: There has been an increased usage of metallic antimicrobial materials to control pathogenic and multi-drug resistant bacteria. Yet, there is a corresponding need to know if this usage leads to genetic adaptations that could produce more harmful strains.
Methodology: Experimental evolution was used to adapt Escherichia coli K-12 MG1655 to excess iron (II) with subsequent genomic analysis. Phenotypic assays and gene expression studies were conducted to demonstrate pleiotropic effects associated with this adaptation and to elucidate potential cellular responses.
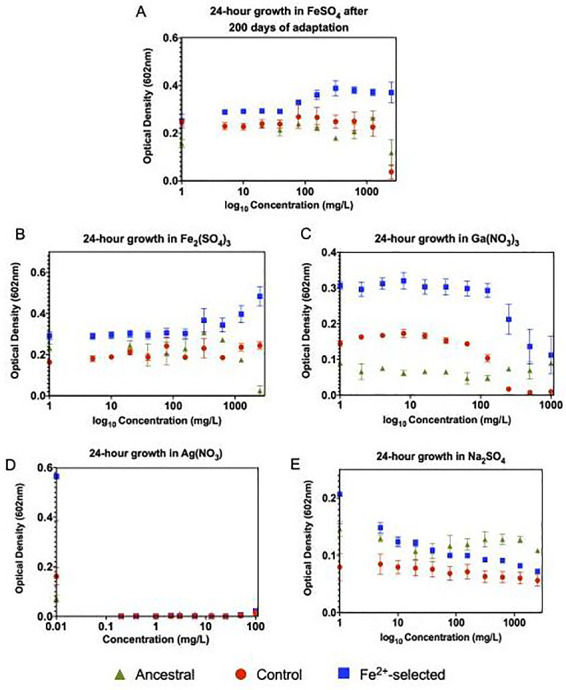
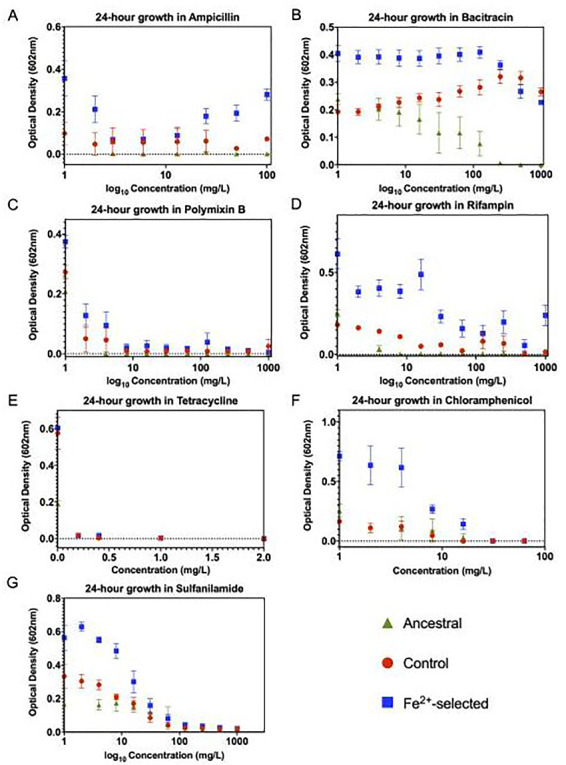
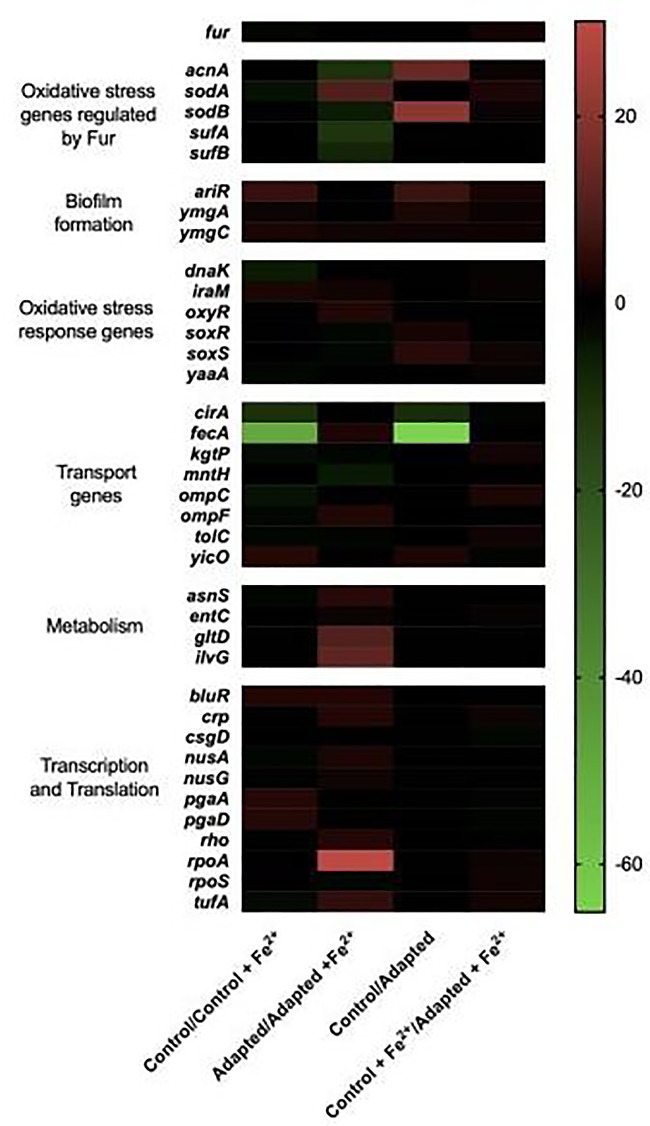
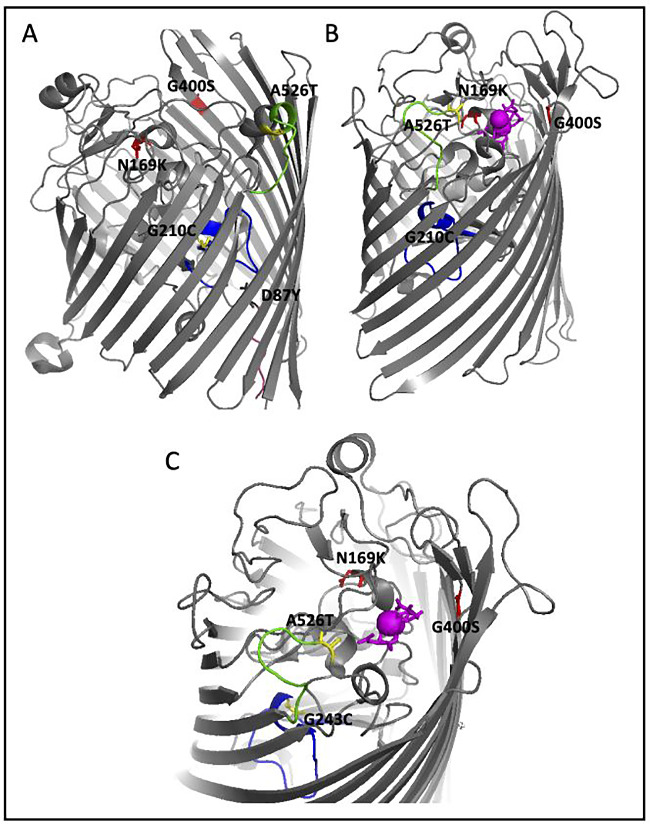
Results: After 200 days of adaptation, populations cultured in excess iron (II), showed a significant increase in 24-h optical densities compared to controls. Furthermore, these populations showed increased resistance toward other metals [iron (III) and gallium (III)] and to traditional antibiotics (bacitracin, rifampin, chloramphenicol and sulfanilamide). Genomic analysis identified selective sweeps in three genes; fecA, ptsP and ilvG unique to the iron (II) resistant populations, and gene expression studies demonstrated that their cellular response may be to downregulate genes involved in iron transport (cirA and fecA) while increasing the oxidative stress response (oxyR, soxS and soxR) prior to FeSO4 exposure.
Conclusions and implications: Together, this indicates that the selected populations can quickly adapt to stressful levels of iron (II). This study is unique in that it demonstrates that E. coli can adapt to environments that contain excess levels of an essential micronutrient while also demonstrating the genomic foundations of the response and the pleiotropic consequences. The fact that adaptation to excess iron also causes increases in general antibiotic resistance is a serious concern. Lay summary: The evolution of iron resistance in E. coli leads to multi-drug and general metal resistance through the acquisition of mutations in three genes (fecA, ptsP and ilvG) while also initiating cellular defenses as part of their normal growth process.
Keywords: E. coli; experimental evolution; gene expression; iron resistance; pleiotropy.
© The Author(s) 2021. Published by Oxford University Press on behalf of the Foundation for Evolution, Medicine, and Public Health.
Figures
References
-
- Colvin RA, Holmes WR, Fontaine CP, Maret W. Cytosolic zinc buffering and muffling: their role in intracellular zinc homeostasis. Metallomics 2010;2:306–17. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
