

Interesting Biochemistries in the Structure and Function of Bacterial Effectors
- PMID: 33718265
- PMCID: PMC7943720
- DOI: 10.3389/fcimb.2021.608860
Interesting Biochemistries in the Structure and Function of Bacterial Effectors
Abstract
Bacterial effector proteins, delivered into host cells by specialized multiprotein secretion systems, are a key mediator of bacterial pathogenesis. Following delivery, they modulate a range of host cellular processes and functions. Strong selective pressures have resulted in bacterial effectors evolving unique structures that can mimic host protein biochemical activity or enable novel and distinct biochemistries. Despite the protein structure-function paradigm, effectors from different bacterial species that share biochemical activities, such as the conjugation of ubiquitin to a substrate, do not necessarily share structural or sequence homology to each other or the eukaryotic proteins that carry out the same function. Furthermore, some bacterial effectors have evolved structural variations to known protein folds which enable different or additional biochemical and physiological functions. Despite the overall low occurrence of intrinsically disordered proteins or regions in prokaryotic proteomes compared to eukaryotes proteomes, bacterial effectors appear to have adopted intrinsically disordered regions that mimic the disordered regions of eukaryotic signaling proteins. In this review, we explore examples of the diverse biochemical properties found in bacterial effectors that enable effector-mediated interference of eukaryotic signaling pathways and ultimately support pathogenesis. Despite challenges in the structural and functional characterisation of effectors, recent progress has been made in understanding the often unusual and fascinating ways in which these virulence factors promote pathogenesis. Nevertheless, continued work is essential to reveal the array of remarkable activities displayed by effectors.
Keywords: bacterial effectors; host-pathogen; pathogenesis; protein organization; secretion systems; structure-function.
Copyright © 2021 Mak and Thurston.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
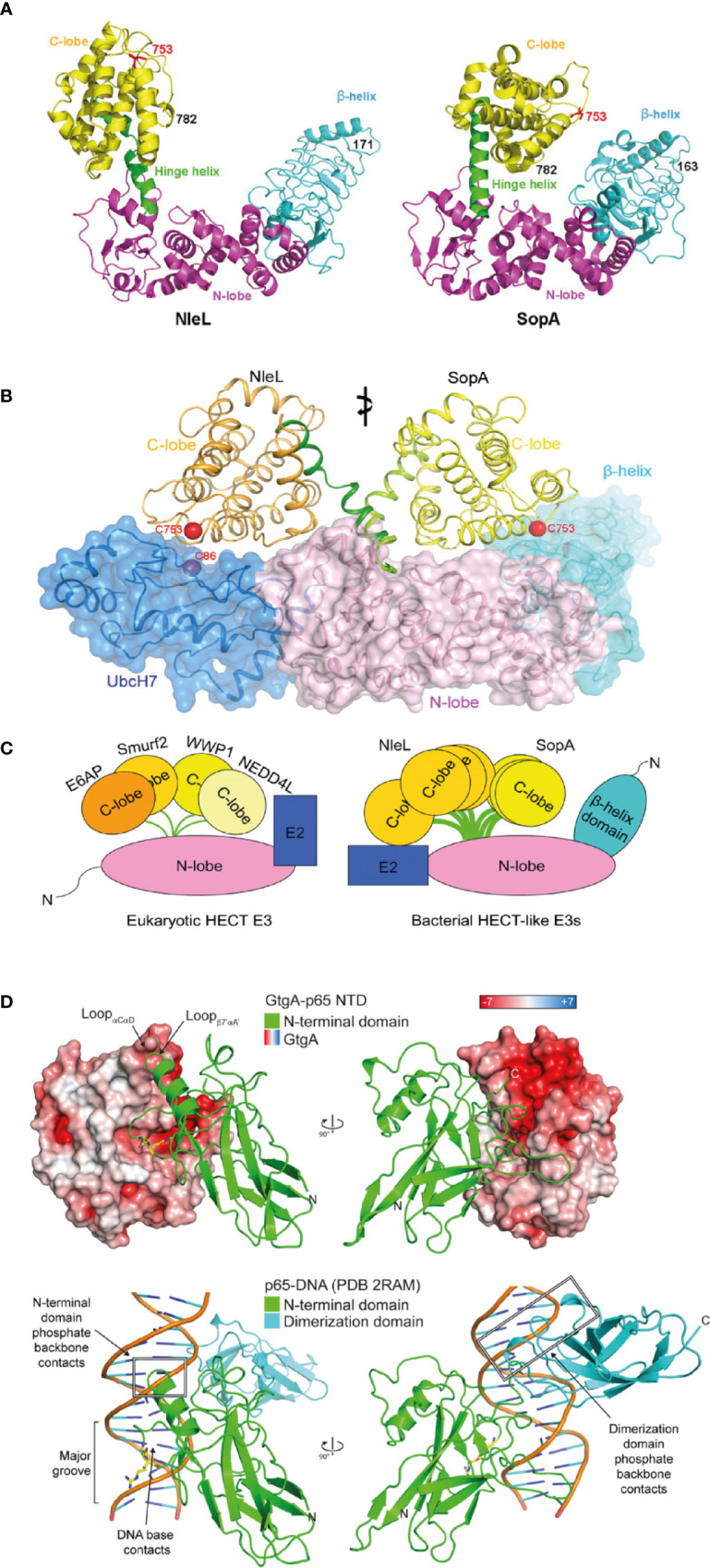
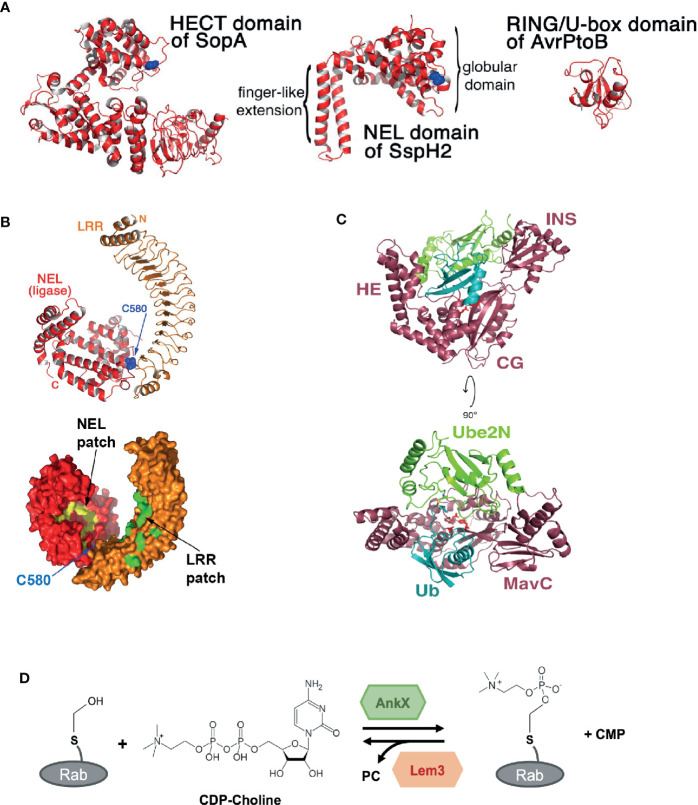
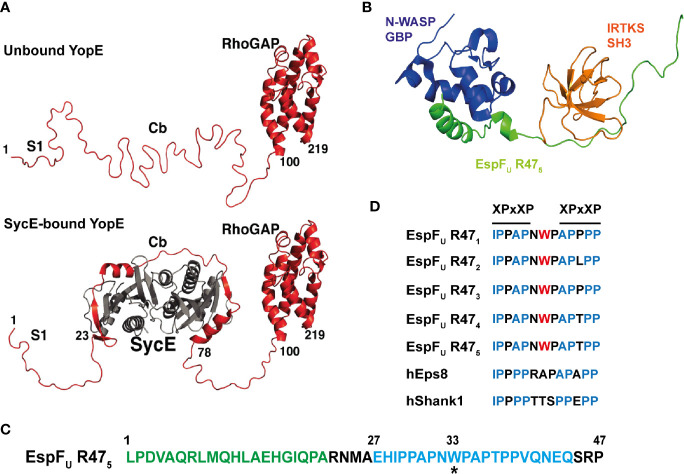
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources