

Functional Mapping of AGO-Associated Zika Virus-Derived Small Interfering RNAs in Neural Stem Cells
- PMID: 33718276
- PMCID: PMC7946837
- DOI: 10.3389/fcimb.2021.628887
Functional Mapping of AGO-Associated Zika Virus-Derived Small Interfering RNAs in Neural Stem Cells
Abstract
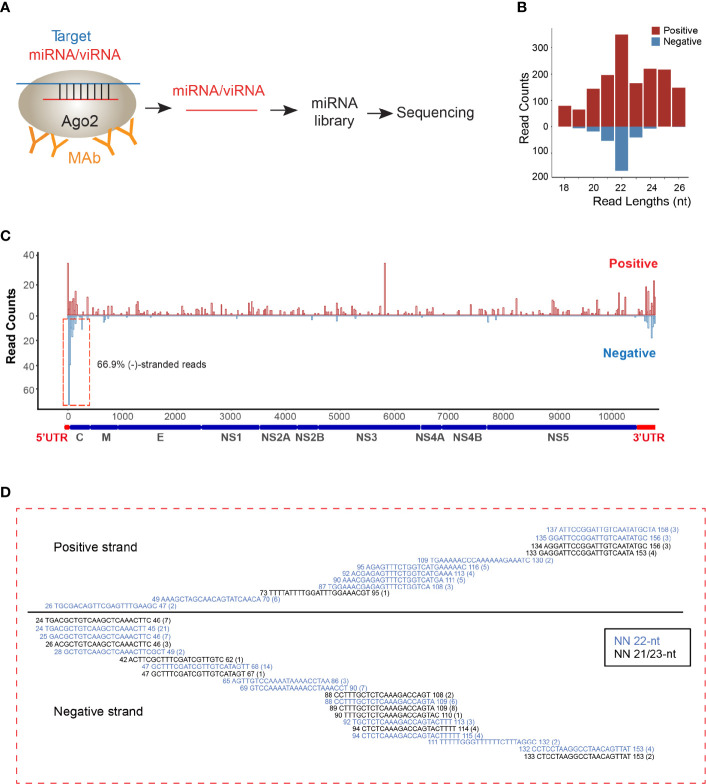
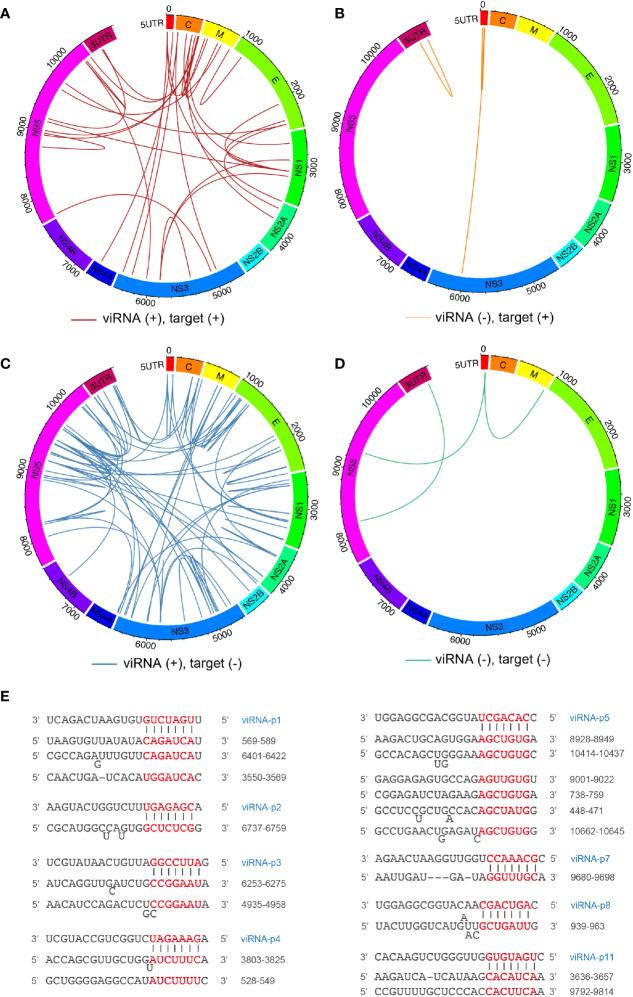
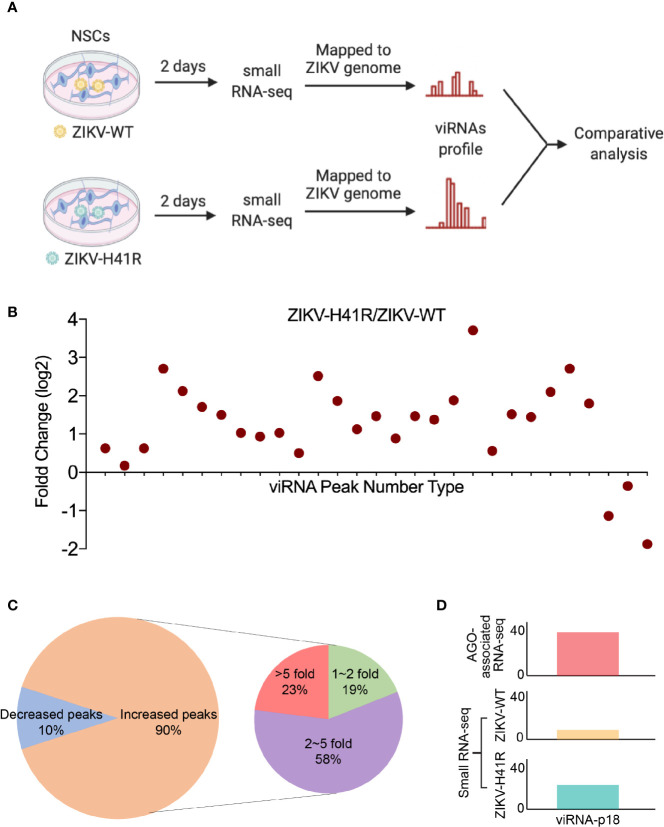
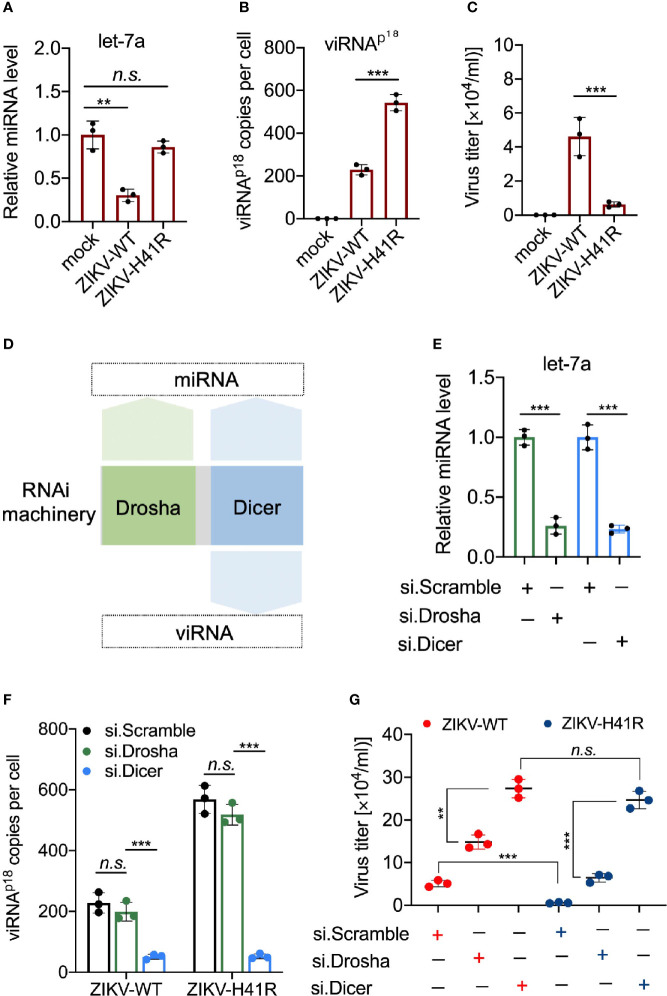
Viral interfering RNA (viRNA) has been identified from several viral genomes via directly deep RNA sequencing of the virus-infected cells, including zika virus (ZIKV). Once produced by endoribonuclease Dicer, viRNAs are loaded onto the Argonaute (AGO) family proteins of the RNA-induced silencing complexes (RISCs) to pair with their RNA targets and initiate the cleavage of target genes. However, the identities of functional ZIKV viRNAs and their viral RNA targets remain largely unknown. Our recent study has shown that ZIKV capsid protein interacted with Dicer and antagonized its endoribonuclease activity, which requires its histidine residue at the 41st amino acid. Accordingly, the engineered ZIKV-H41R loss-of-function (LOF) mutant virus no longer suppresses Dicer enzymatic activity nor inhibits miRNA biogenesis in NSCs. By combining AGO-associated RNA sequencing, deep sequencing analysis in ZIKV-infected human neural stem cells (NSCs), and miRanda target scanning, we defined 29 ZIKV derived viRNA profiles in NSCs, and established a complex interaction network between the viRNAs and their viral targets. More importantly, we found that viRNA production from the ZIKV mRNA is dependent on Dicer function and is a limiting factor for ZIKV virulence in NSCs. As a result, much higher levels of viRNAs generated from the ZIKV-H41R virus-infected NSCs. Therefore, our mapping of viRNAs to their RNA targets paves a way to further investigate how viRNAs play the role in anti-viral mechanisms, and perhaps other unknown biological functions.
Keywords: Dicer; RNA interference (RNAi); neural stem cells (NSCs); viral interfering RNA (viRNA); zika virus (ZIKV).
Copyright © 2021 Zeng, Luo, Dong, Xie, Liang, Yan, Liang and Zhao.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
The Zika Virus Capsid Disrupts Corticogenesis by Suppressing Dicer Activity and miRNA Biogenesis.Cell Stem Cell. 2020 Oct 1;27(4):618-632.e9. doi: 10.1016/j.stem.2020.07.012. Epub 2020 Aug 6. Cell Stem Cell. 2020. PMID: 32763144 Free PMC article.
-
The subgenomic flaviviral RNA suppresses RNA interference through competing with siRNAs for binding RISC components.J Virol. 2024 Feb 20;98(2):e0195423. doi: 10.1128/jvi.01954-23. Epub 2024 Jan 30. J Virol. 2024. PMID: 38289102 Free PMC article.
-
Antiviral RNA interference responses induced by Semliki Forest virus infection of mosquito cells: characterization, origin, and frequency-dependent functions of virus-derived small interfering RNAs.J Virol. 2011 Mar;85(6):2907-17. doi: 10.1128/JVI.02052-10. Epub 2010 Dec 29. J Virol. 2011. PMID: 21191029 Free PMC article.
-
Functional RNA during Zika virus infection.Virus Res. 2018 Aug 2;254:41-53. doi: 10.1016/j.virusres.2017.08.015. Epub 2017 Aug 31. Virus Res. 2018. PMID: 28864425 Review.
-
Neuroimmune Evasion of Zika Virus to Facilitate Viral Pathogenesis.Front Cell Infect Microbiol. 2021 Oct 26;11:662447. doi: 10.3389/fcimb.2021.662447. eCollection 2021. Front Cell Infect Microbiol. 2021. PMID: 34765564 Free PMC article. Review.
Cited by
-
Identification of antiviral RNAi regulators, ILF3/DHX9, recruit at ZIKV stem loop B to protect against ZIKV induced microcephaly.Nat Commun. 2025 Feb 26;16(1):1991. doi: 10.1038/s41467-025-56859-x. Nat Commun. 2025. PMID: 40011444 Free PMC article.
-
MicroRNA let-7 and viral infections: focus on mechanisms of action.Cell Mol Biol Lett. 2022 Feb 14;27(1):14. doi: 10.1186/s11658-022-00317-9. Cell Mol Biol Lett. 2022. PMID: 35164678 Free PMC article. Review.
-
Decoding protein-RNA interactions using CLIP-based methodologies.Nat Rev Genet. 2024 Dec;25(12):879-895. doi: 10.1038/s41576-024-00749-3. Epub 2024 Jul 9. Nat Rev Genet. 2024. PMID: 38982239 Review.
-
Mammalian viral suppressors of RNA interference.Trends Biochem Sci. 2022 Nov;47(11):978-988. doi: 10.1016/j.tibs.2022.05.001. Epub 2022 May 23. Trends Biochem Sci. 2022. PMID: 35618579 Free PMC article. Review.
-
SARS-CoV-2 RNAs are processed into 22-nt vsRNAs in Vero cells.Front Immunol. 2022 Oct 28;13:1008084. doi: 10.3389/fimmu.2022.1008084. eCollection 2022. Front Immunol. 2022. PMID: 36389816 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials