

GAF is essential for zygotic genome activation and chromatin accessibility in the early Drosophila embryo
- PMID: 33720012
- PMCID: PMC8079149
- DOI: 10.7554/eLife.66668
GAF is essential for zygotic genome activation and chromatin accessibility in the early Drosophila embryo
Abstract
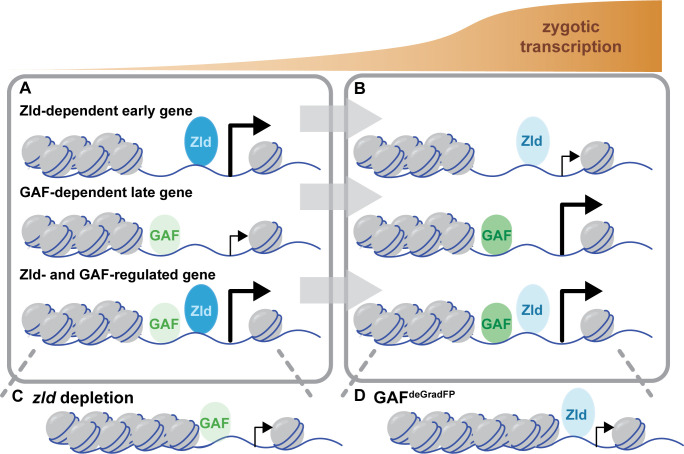
Following fertilization, the genomes of the germ cells are reprogrammed to form the totipotent embryo. Pioneer transcription factors are essential for remodeling the chromatin and driving the initial wave of zygotic gene expression. In Drosophila melanogaster, the pioneer factor Zelda is essential for development through this dramatic period of reprogramming, known as the maternal-to-zygotic transition (MZT). However, it was unknown whether additional pioneer factors were required for this transition. We identified an additional maternally encoded factor required for development through the MZT, GAGA Factor (GAF). GAF is necessary to activate widespread zygotic transcription and to remodel the chromatin accessibility landscape. We demonstrated that Zelda preferentially controls expression of the earliest transcribed genes, while genes expressed during widespread activation are predominantly dependent on GAF. Thus, progression through the MZT requires coordination of multiple pioneer-like factors, and we propose that as development proceeds control is gradually transferred from Zelda to GAF.
Keywords: D. melanogaster; chromatin; chromosomes; gene expression; maternal-to-zygotic transition; pioneer factor; transcription; zygotic genome activation.
Plain language summary
Most cells in an organism share the exact same genetic information, yet they still adopt distinct identities. This diversity emerges because only a selection of genes is switched on at any given time in a cell. Proteins that latch onto DNA control this specificity by activating certain genes at the right time. However, to perform this role they first need to physically access DNA: this can be difficult as the genetic information is tightly compacted so it can fit in a cell. A group of proteins can help to unpack the genome to uncover the genes that can then be accessed and activated. While these ‘pioneer factors’ can therefore shape the identity of a cell, much remains unknown about how they can work together to do so. For instance, the pioneer factor Zelda is essential in early fruit fly development, as it enables the genetic information of the egg and sperm to undergo dramatic reprogramming and generate a new organism. Yet, it was unclear whether additional helpers were required for this transition. Using this animal system, Gaskill, Gibson et al. identified GAGA Factor as a protein which works with Zelda to open up and reprogram hundreds of different sections along the genome of fruit fly embryos. This tag-team effort started with Zelda being important initially to activate genes; regulation was then handed over for GAGA Factor to continue the process. Without either protein, the embryo died. Getting a glimpse into early genetic events during fly development provides insights that are often applicable to other animals such as fish and mammals. Ultimately, this research may help scientists to understand how things can go wrong in human embryos.
© 2021, Gaskill et al.
Conflict of interest statement
MG, TG, EL, MH No competing interests declared
Figures















Similar articles
-
Genome-wide search for Zelda-like chromatin signatures identifies GAF as a pioneer factor in early fly development.Epigenetics Chromatin. 2017 Jul 4;10(1):33. doi: 10.1186/s13072-017-0141-5. Epigenetics Chromatin. 2017. PMID: 28676122 Free PMC article.
-
Zelda is differentially required for chromatin accessibility, transcription factor binding, and gene expression in the early Drosophila embryo.Genome Res. 2015 Nov;25(11):1715-26. doi: 10.1101/gr.192682.115. Epub 2015 Sep 2. Genome Res. 2015. PMID: 26335634 Free PMC article.
-
Zelda binding in the early Drosophila melanogaster embryo marks regions subsequently activated at the maternal-to-zygotic transition.PLoS Genet. 2011 Oct;7(10):e1002266. doi: 10.1371/journal.pgen.1002266. Epub 2011 Oct 20. PLoS Genet. 2011. PMID: 22028662 Free PMC article.
-
Regulatory principles governing the maternal-to-zygotic transition: insights from Drosophila melanogaster.Open Biol. 2018 Dec;8(12):180183. doi: 10.1098/rsob.180183. Open Biol. 2018. PMID: 30977698 Free PMC article. Review.
-
Epigenetic inheritance and gene expression regulation in early Drosophila embryos.EMBO Rep. 2024 Oct;25(10):4131-4152. doi: 10.1038/s44319-024-00245-z. Epub 2024 Sep 16. EMBO Rep. 2024. PMID: 39285248 Free PMC article. Review.
Cited by
-
The Drosophila embryo as a tabula rasa for the epigenome.Fac Rev. 2022 Dec 23;11:40. doi: 10.12703/r/11-40. eCollection 2022. Fac Rev. 2022. PMID: 36644296 Free PMC article. Review.
-
Hybridization led to a rewired pluripotency network in the allotetraploid Xenopus laevis.Elife. 2023 Oct 3;12:e83952. doi: 10.7554/eLife.83952. Elife. 2023. PMID: 37787392 Free PMC article.
-
Setting the stage for development: the maternal-to-zygotic transition in Drosophila.Genetics. 2023 Oct 4;225(2):iyad142. doi: 10.1093/genetics/iyad142. Genetics. 2023. PMID: 37616526 Free PMC article. Review.
-
Tissue-specific RNA Polymerase II promoter-proximal pause release and burst kinetics in a Drosophila embryonic patterning network.Genome Biol. 2024 Jan 2;25(1):2. doi: 10.1186/s13059-023-03135-0. Genome Biol. 2024. PMID: 38166964 Free PMC article.
-
Stepwise modifications of transcriptional hubs link pioneer factor activity to a burst of transcription.Nat Commun. 2023 Aug 10;14(1):4848. doi: 10.1038/s41467-023-40485-6. Nat Commun. 2023. PMID: 37563108 Free PMC article.
References
-
- Bhat KM, Farkas G, Karch F, Gyurkovics H, Gausz J, Schedl P. The GAGA factor is required in the early Drosophila embryo not only for transcriptional regulation but also for nuclear division. Development. 1996;122:1113–1124. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
