

Directed Evolution of an Enhanced POU Reprogramming Factor for Cell Fate Engineering
- PMID: 33720298
- PMCID: PMC8233511
- DOI: 10.1093/molbev/msab075
Directed Evolution of an Enhanced POU Reprogramming Factor for Cell Fate Engineering
Abstract
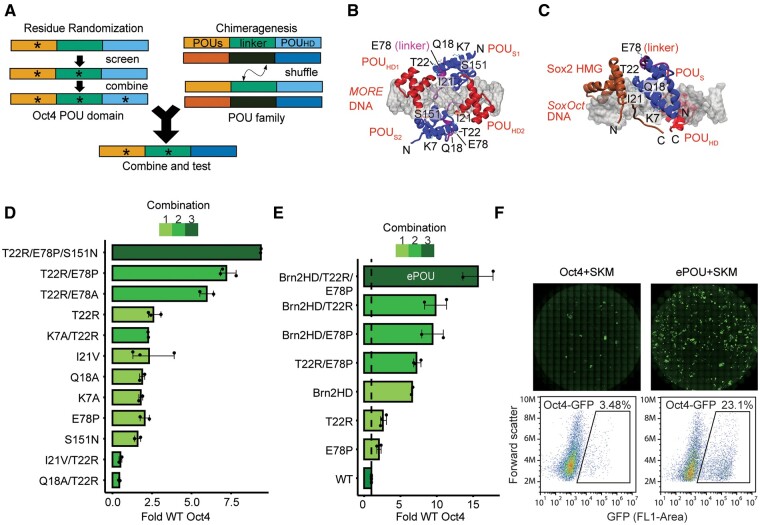
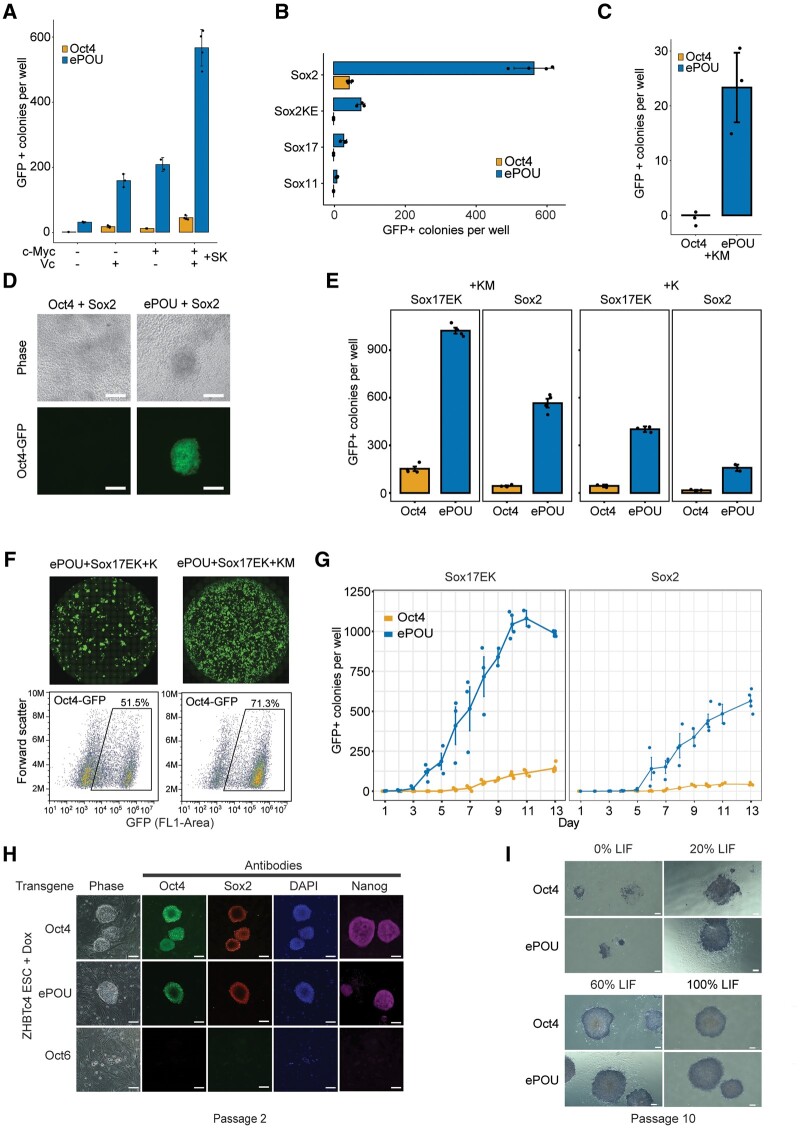
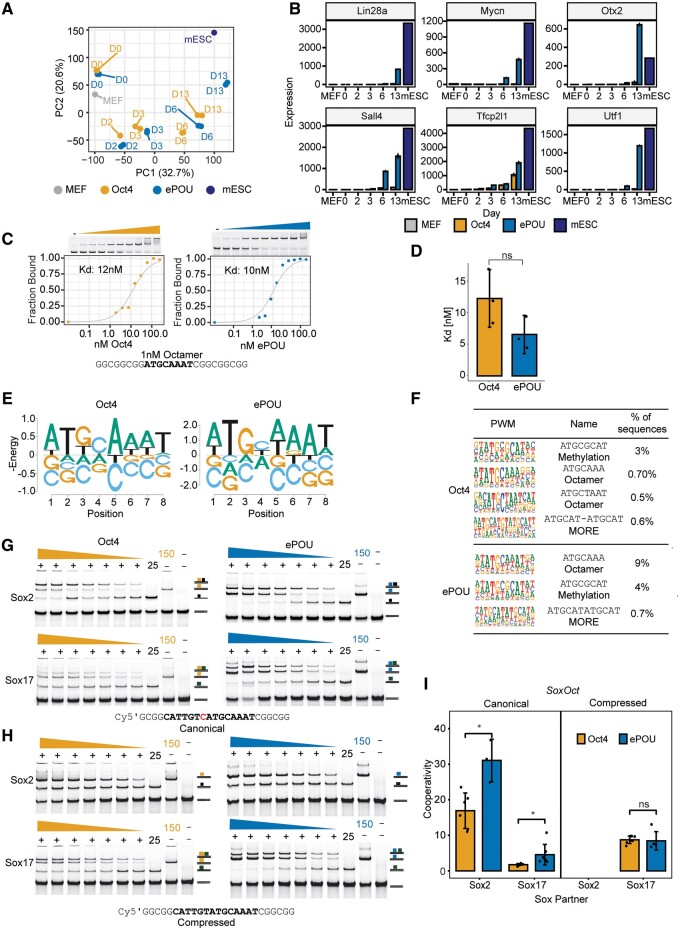
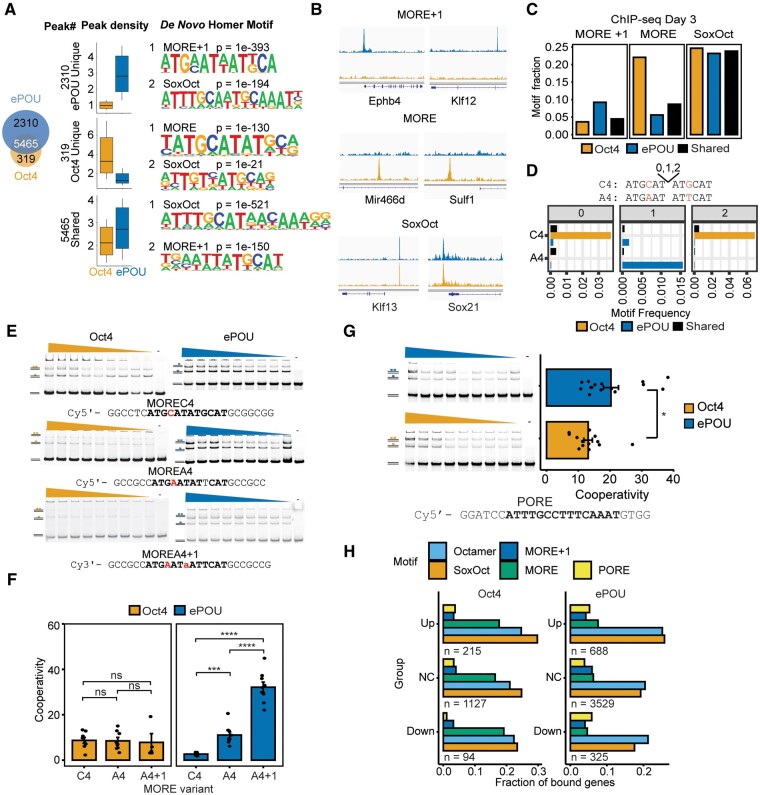
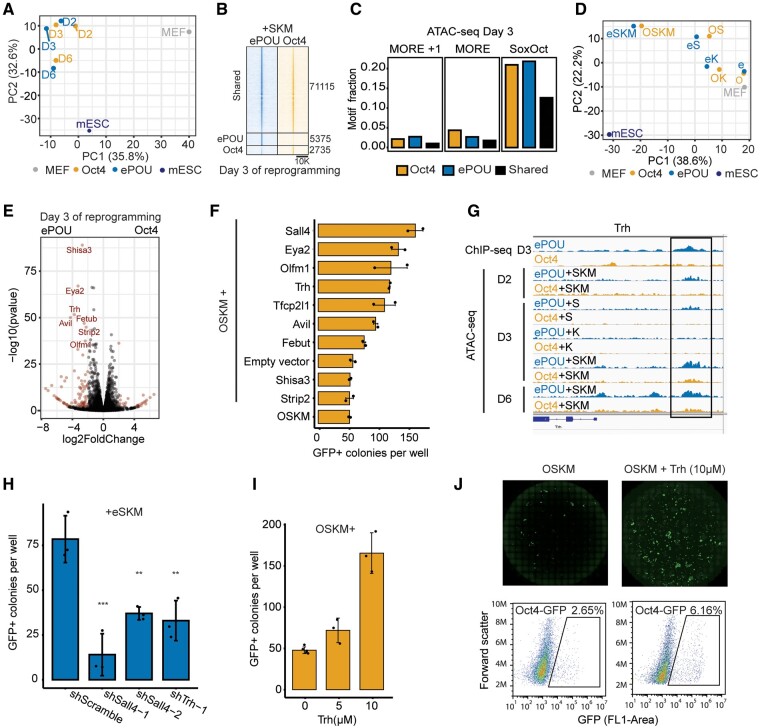
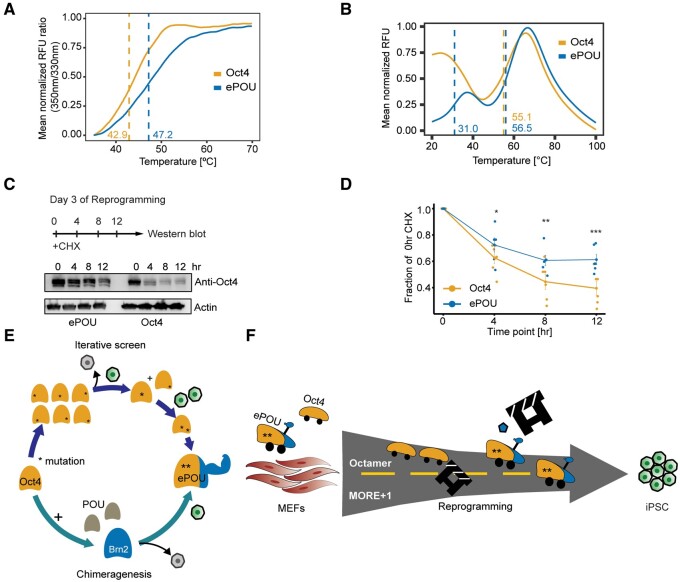
Transcription factor-driven cell fate engineering in pluripotency induction, transdifferentiation, and forward reprogramming requires efficiency, speed, and maturity for widespread adoption and clinical translation. Here, we used Oct4, Sox2, Klf4, and c-Myc driven pluripotency reprogramming to evaluate methods for enhancing and tailoring cell fate transitions, through directed evolution with iterative screening of pooled mutant libraries and phenotypic selection. We identified an artificially evolved and enhanced POU factor (ePOU) that substantially outperforms wild-type Oct4 in terms of reprogramming speed and efficiency. In contrast to Oct4, not only can ePOU induce pluripotency with Sox2 alone, but it can also do so in the absence of Sox2 in a three-factor ePOU/Klf4/c-Myc cocktail. Biochemical assays combined with genome-wide analyses showed that ePOU possesses a new preference to dimerize on palindromic DNA elements. Yet, the moderate capacity of Oct4 to function as a pioneer factor, its preference to bind octamer DNA and its capability to dimerize with Sox2 and Sox17 proteins remain unchanged in ePOU. Compared with Oct4, ePOU is thermodynamically stabilized and persists longer in reprogramming cells. In consequence, ePOU: 1) differentially activates several genes hitherto not implicated in reprogramming, 2) reveals an unappreciated role of thyrotropin-releasing hormone signaling, and 3) binds a distinct class of retrotransposons. Collectively, these features enable ePOU to accelerate the establishment of the pluripotency network. This demonstrates that the phenotypic selection of novel factor variants from mammalian cells with desired properties is key to advancing cell fate conversions with artificially evolved biomolecules.
Keywords: POU; cell fate conversion; molecular evolution; protein engineering; reprogramming; transcription factor.
© The Author(s) 2021. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures
References
-
- Aksoy I, Jauch R, Eras V, Chng WB, Chen J, Divakar U, Ng CK, Kolatkar PR, Stanton LW.. 2013. Sox transcription factors require selective interactions with oct4 and specific transactivation functions to mediate reprogramming. Stem Cells 31(12):2632–2646. - PubMed
-
- Asial I, Cheng YX, Engman H, Dollhopf M, Wu B, Nordlund P, Cornvik T.. 2013. Engineering protein thermostability using a generic activity-independent biophysical screen inside the cell. Nat Commun. 4:2901. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
