

The velvet protein Vel1 controls initial plant root colonization and conidia formation for xylem distribution in Verticillium wilt
- PMID: 33720931
- PMCID: PMC7993770
- DOI: 10.1371/journal.pgen.1009434
The velvet protein Vel1 controls initial plant root colonization and conidia formation for xylem distribution in Verticillium wilt
Abstract
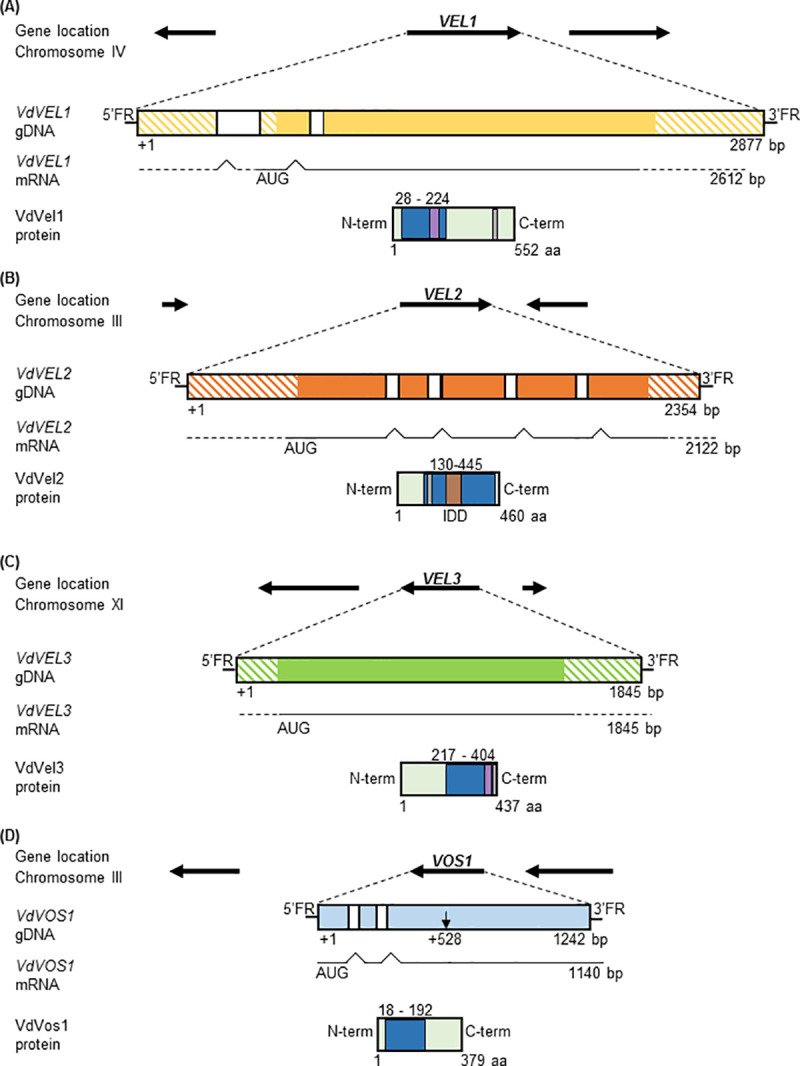
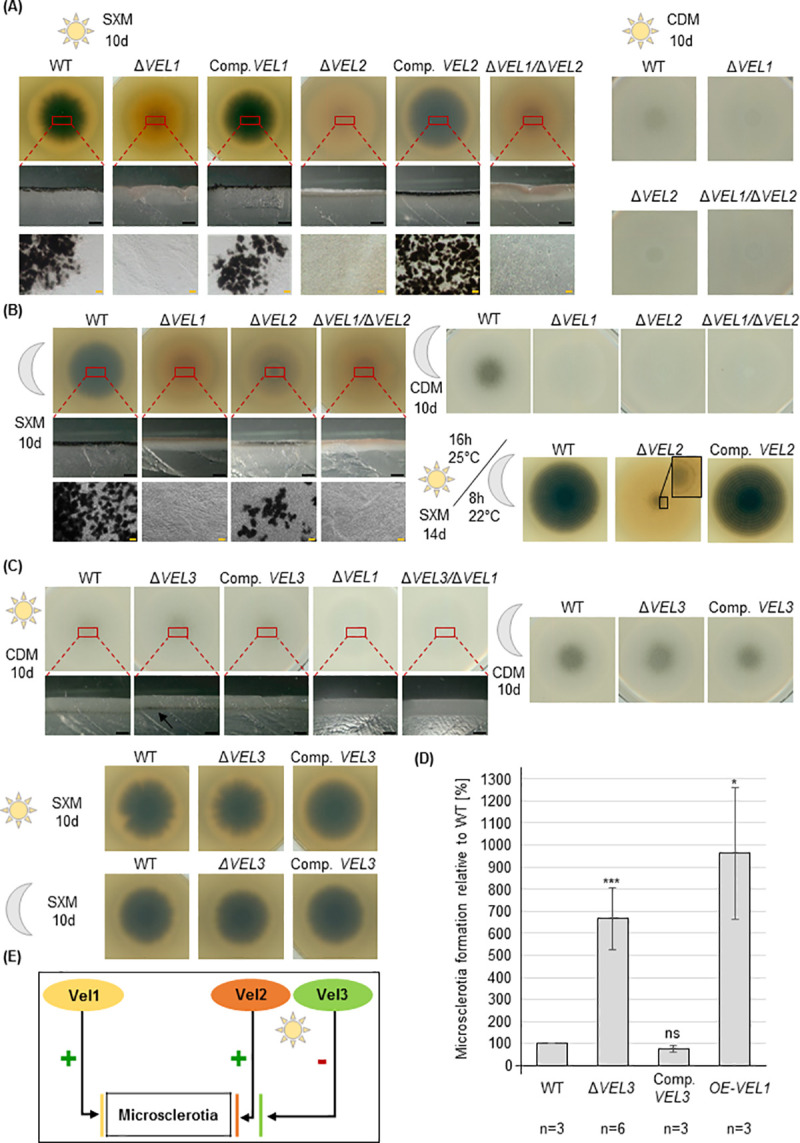
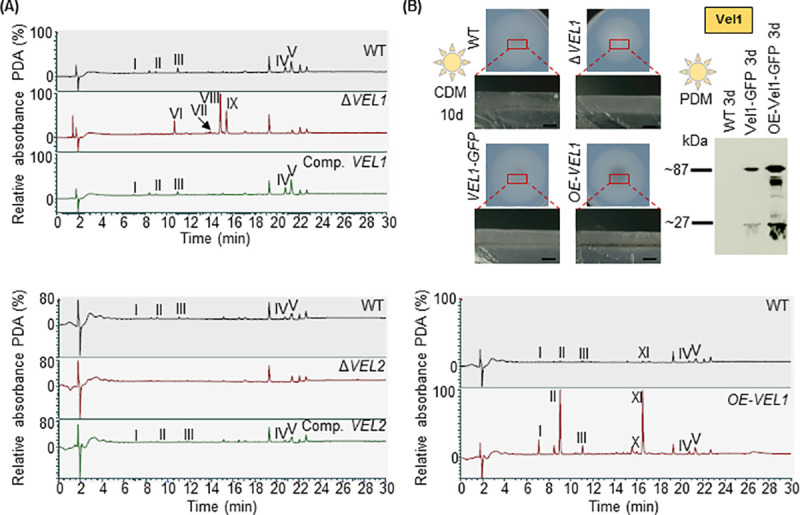
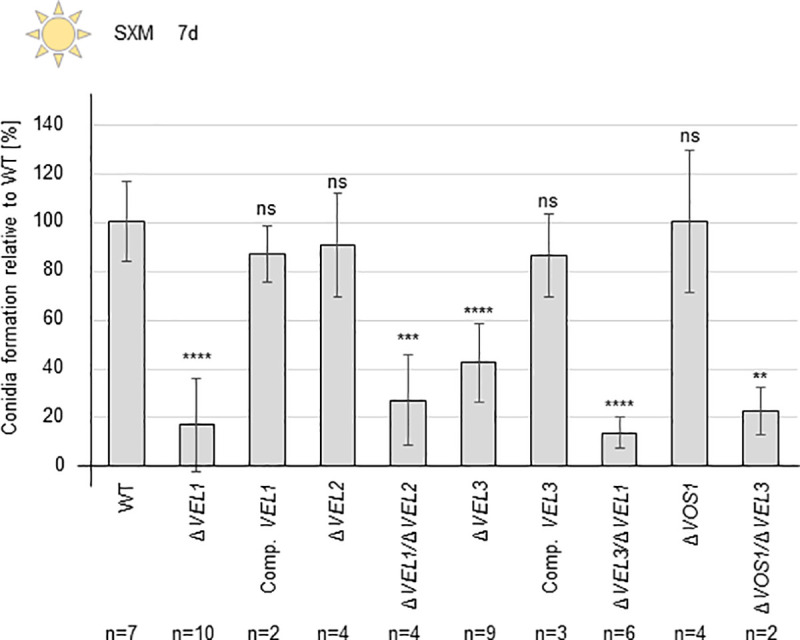
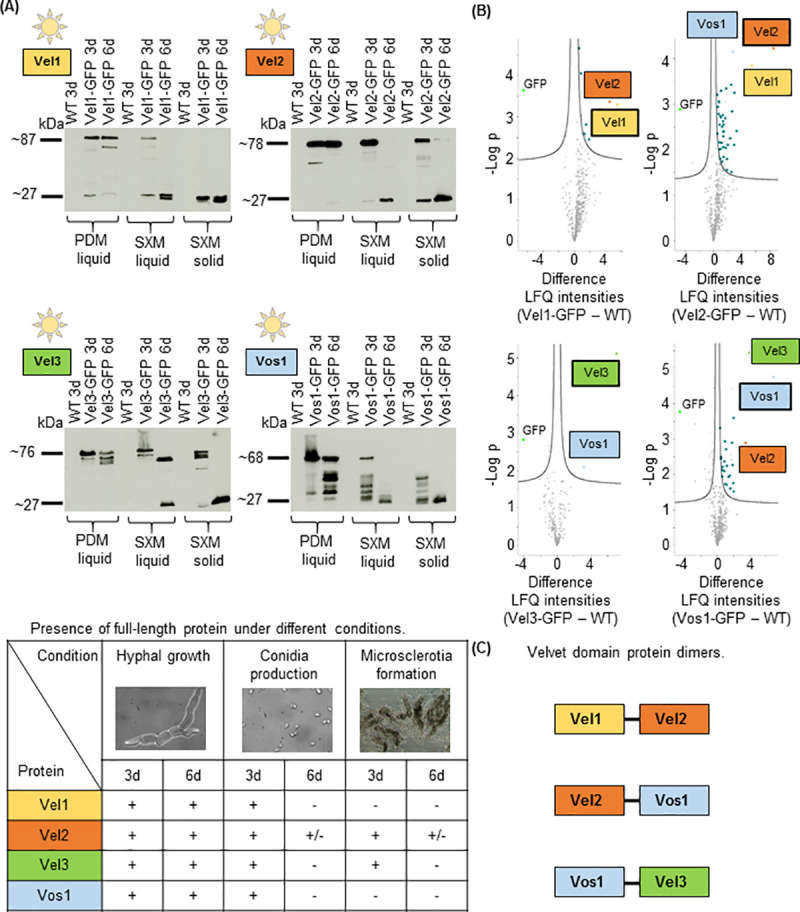
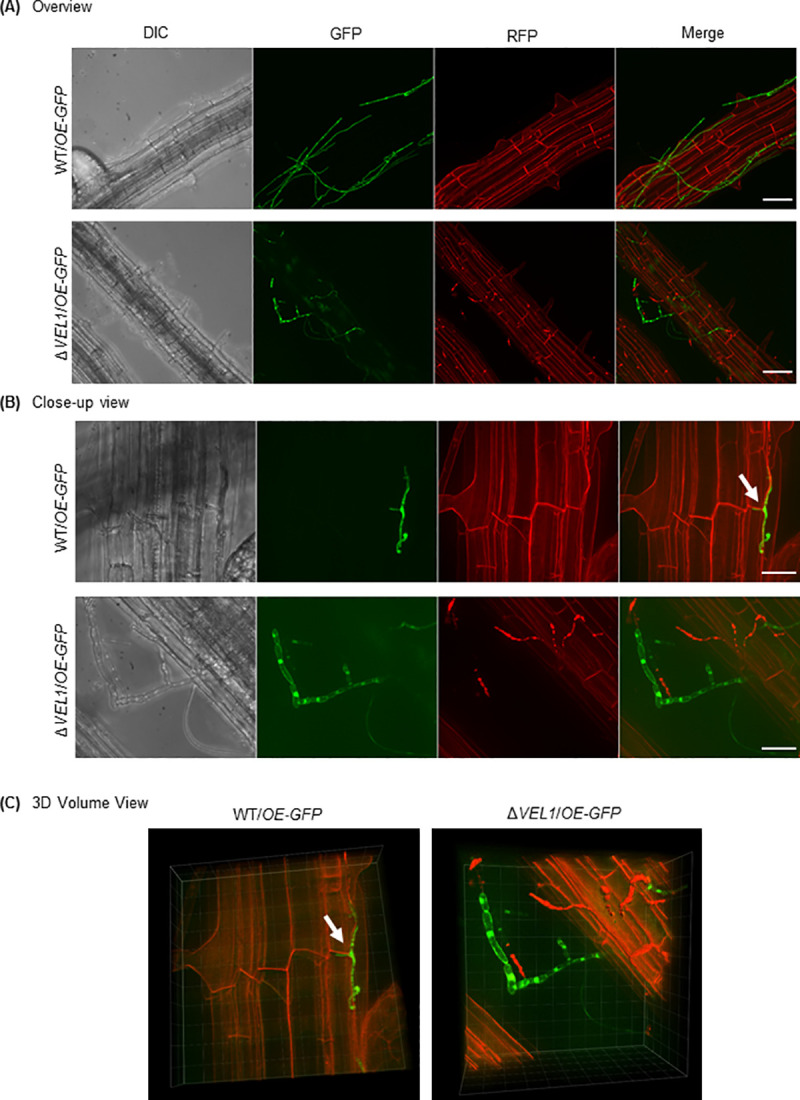
The conserved fungal velvet family regulatory proteins link development and secondary metabolite production. The velvet domain for DNA binding and dimerization is similar to the structure of the Rel homology domain of the mammalian NF-κB transcription factor. A comprehensive study addressed the functions of all four homologs of velvet domain encoding genes in the fungal life cycle of the soil-borne plant pathogenic fungus Verticillium dahliae. Genetic, cell biological, proteomic and metabolomic analyses of Vel1, Vel2, Vel3 and Vos1 were combined with plant pathogenicity experiments. Different phases of fungal growth, development and pathogenicity require V. dahliae velvet proteins, including Vel1-Vel2, Vel2-Vos1 and Vel3-Vos1 heterodimers, which are already present during vegetative hyphal growth. The major novel finding of this study is that Vel1 is necessary for initial plant root colonization and together with Vel3 for propagation in planta by conidiation. Vel1 is needed for disease symptom induction in tomato. Vel1, Vel2, and Vel3 control the formation of microsclerotia in senescent plants. Vel1 is the most important among all four V. dahliae velvet proteins with a wide variety of functions during all phases of the fungal life cycle in as well as ex planta.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures


References
-
- Wilhelm S. Longevity of the Verticillium wilt fungus in the laboratory and field. Phytopathology. 1955;45:180–181.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
