

Exploiting Tumor Neoantigens to Target Cancer Evolution: Current Challenges and Promising Therapeutic Approaches
- PMID: 33722796
- PMCID: PMC8102318
- DOI: 10.1158/2159-8290.CD-20-1575
Exploiting Tumor Neoantigens to Target Cancer Evolution: Current Challenges and Promising Therapeutic Approaches
Abstract
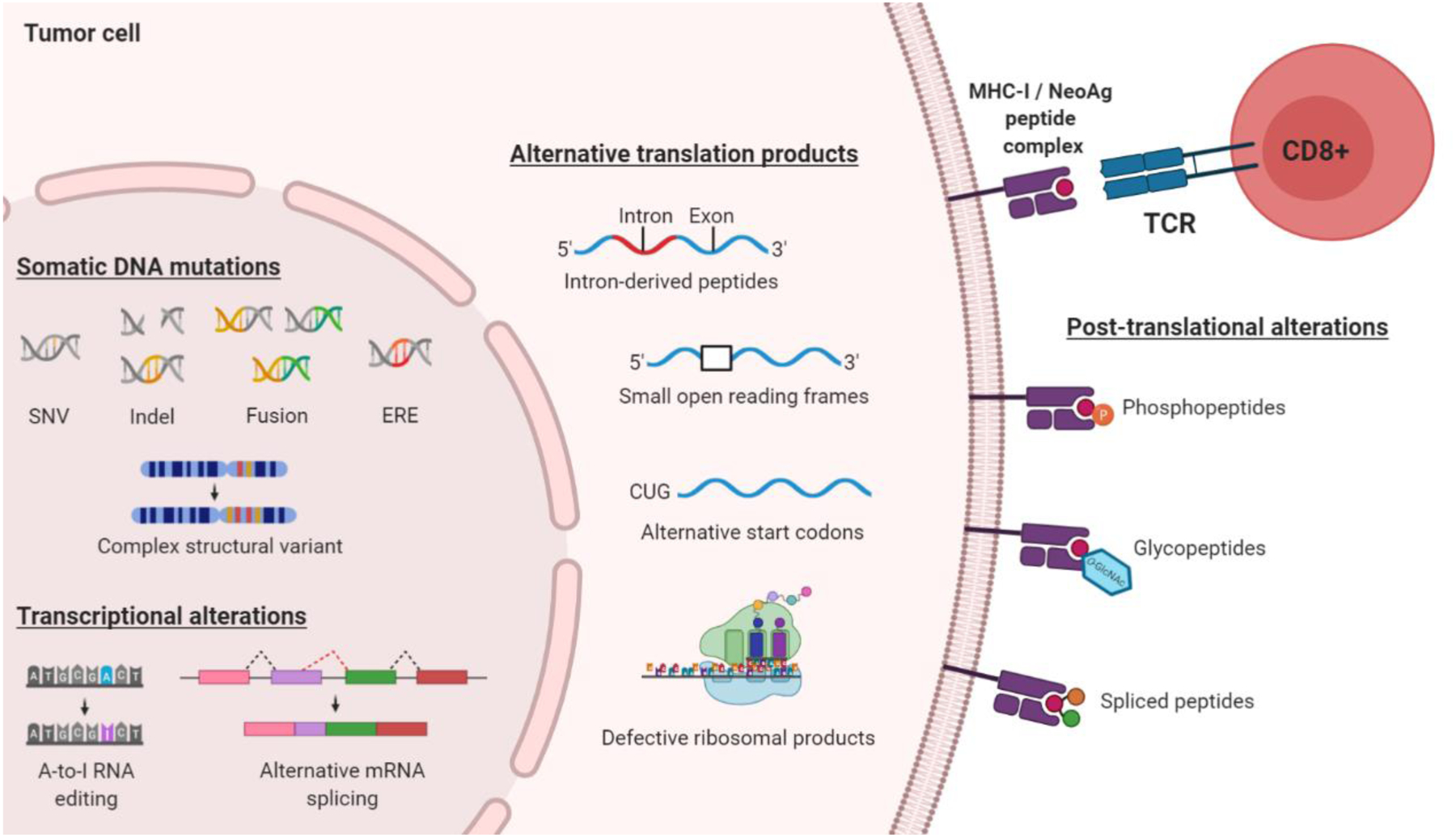
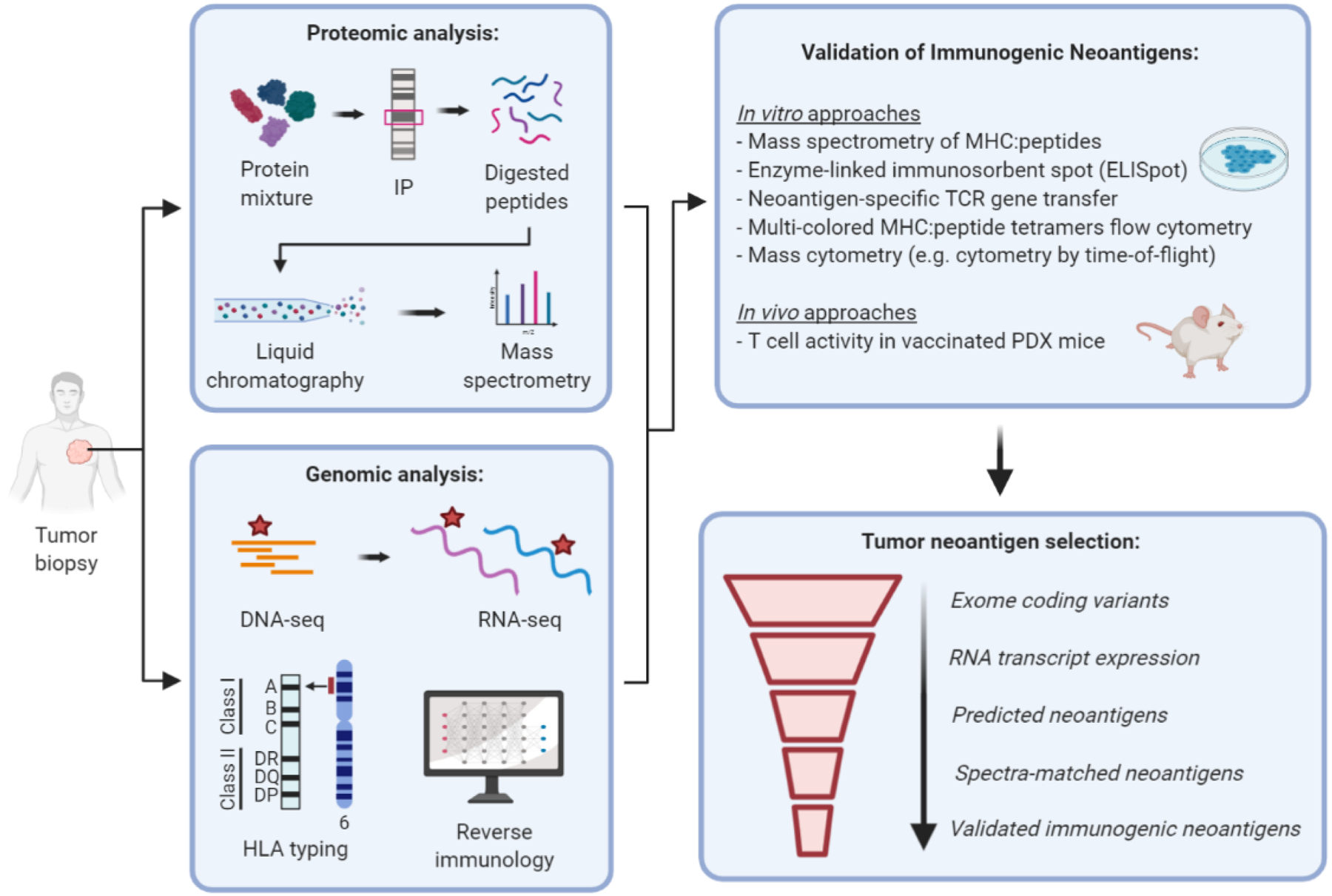
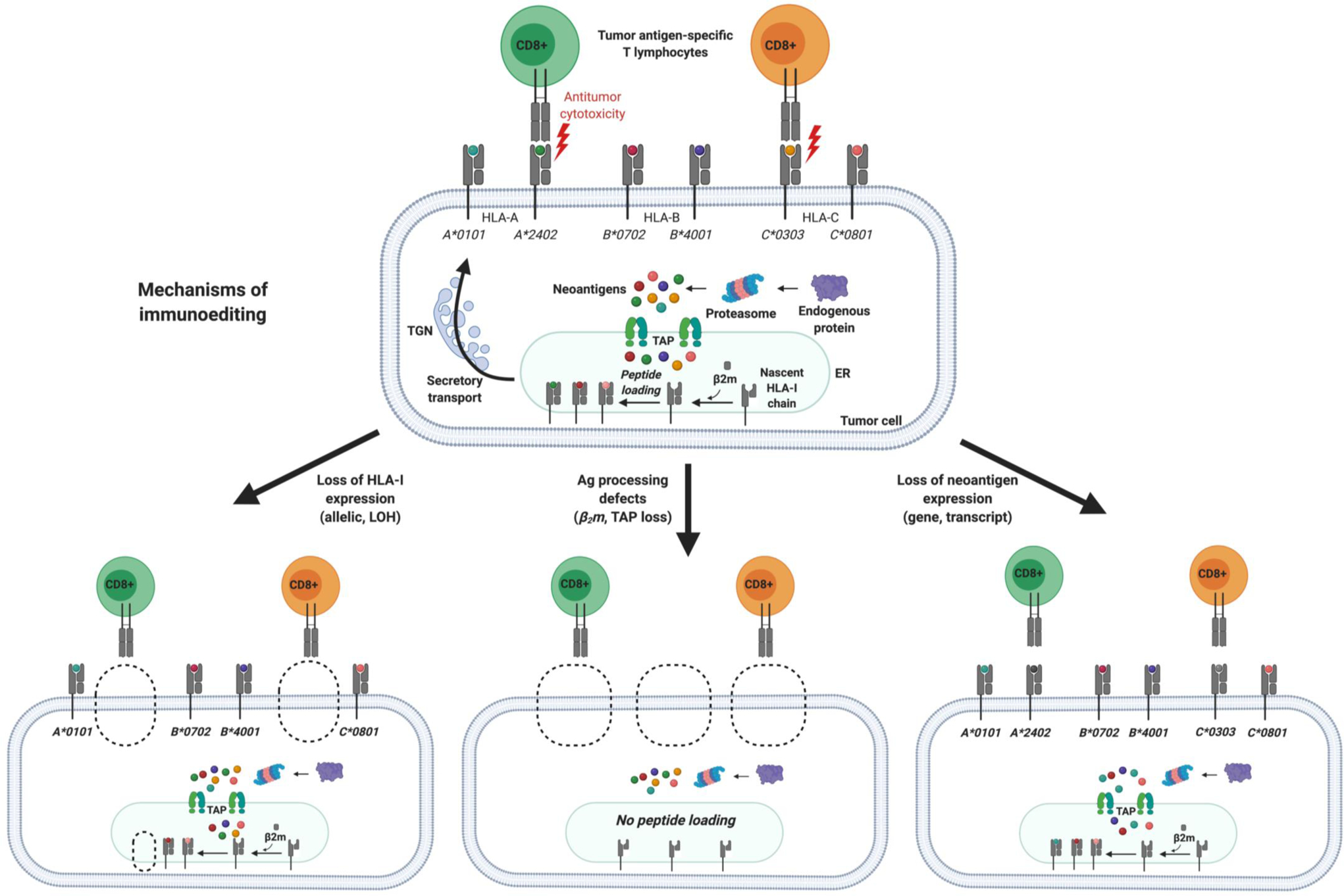
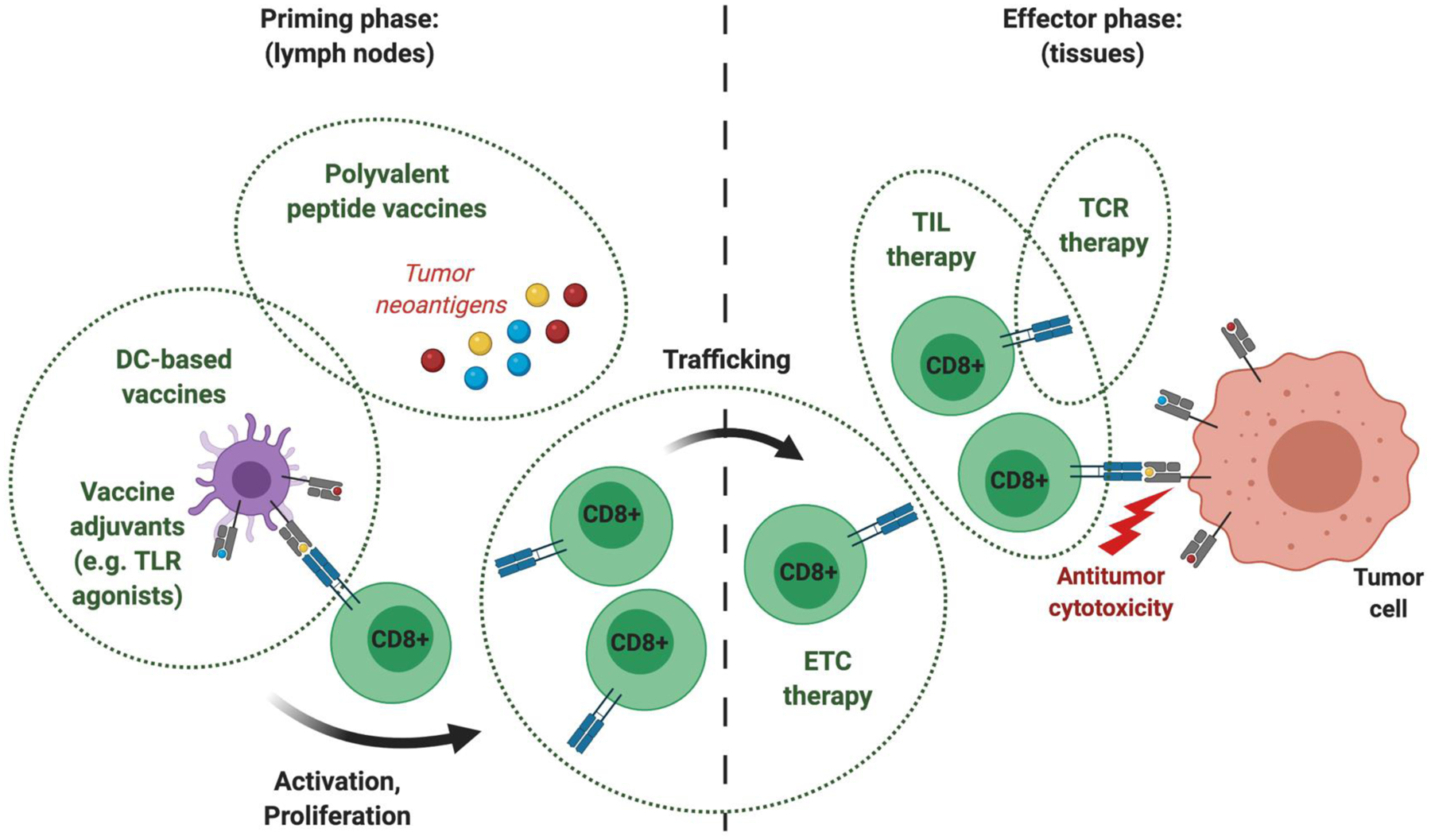
Immunotherapeutic manipulation of the antitumor immune response offers an attractive strategy to target genomic instability in cancer. A subset of tumor-specific somatic mutations can be translated into immunogenic and HLA-bound epitopes called neoantigens, which can induce the activation of helper and cytotoxic T lymphocytes. However, cancer immunoediting and immunosuppressive mechanisms often allow tumors to evade immune recognition. Recent evidence also suggests that the tumor neoantigen landscape extends beyond epitopes originating from nonsynonymous single-nucleotide variants in the coding exome. Here we review emerging approaches for identifying, prioritizing, and immunologically targeting personalized neoantigens using polyvalent cancer vaccines and T-cell receptor gene therapy. SIGNIFICANCE: Several major challenges currently impede the clinical efficacy of neoantigen-directed immunotherapy, such as the relative infrequency of immunogenic neoantigens, suboptimal potency and priming of de novo tumor-specific T cells, and tumor cell-intrinsic and -extrinsic mechanisms of immune evasion. A deeper understanding of these biological barriers could help facilitate the development of effective and durable immunotherapy for any type of cancer, including immunologically "cold" tumors that are otherwise therapeutically resistant.
©2021 American Association for Cancer Research.
Conflict of interest statement
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Figures
References
-
- Fearon ER, Vogelstein B. A genetic model for colorectal tumorigenesis. Cell 1990;61:759–67. - PubMed
-
- Schuh A, Becq J, Humphray S, Alexa A, Burns A, Clifford R, et al. Monitoring chronic lymphocytic leukemia progression by whole genome sequencing reveals heterogeneous clonal evolution patterns. Blood 2012;120:4191–6. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
