

Chromatin remodelers and lineage-specific factors interact to target enhancers to establish proneurosensory fate within otic ectoderm
- PMID: 33723076
- PMCID: PMC8000026
- DOI: 10.1073/pnas.2025196118
Chromatin remodelers and lineage-specific factors interact to target enhancers to establish proneurosensory fate within otic ectoderm
Abstract
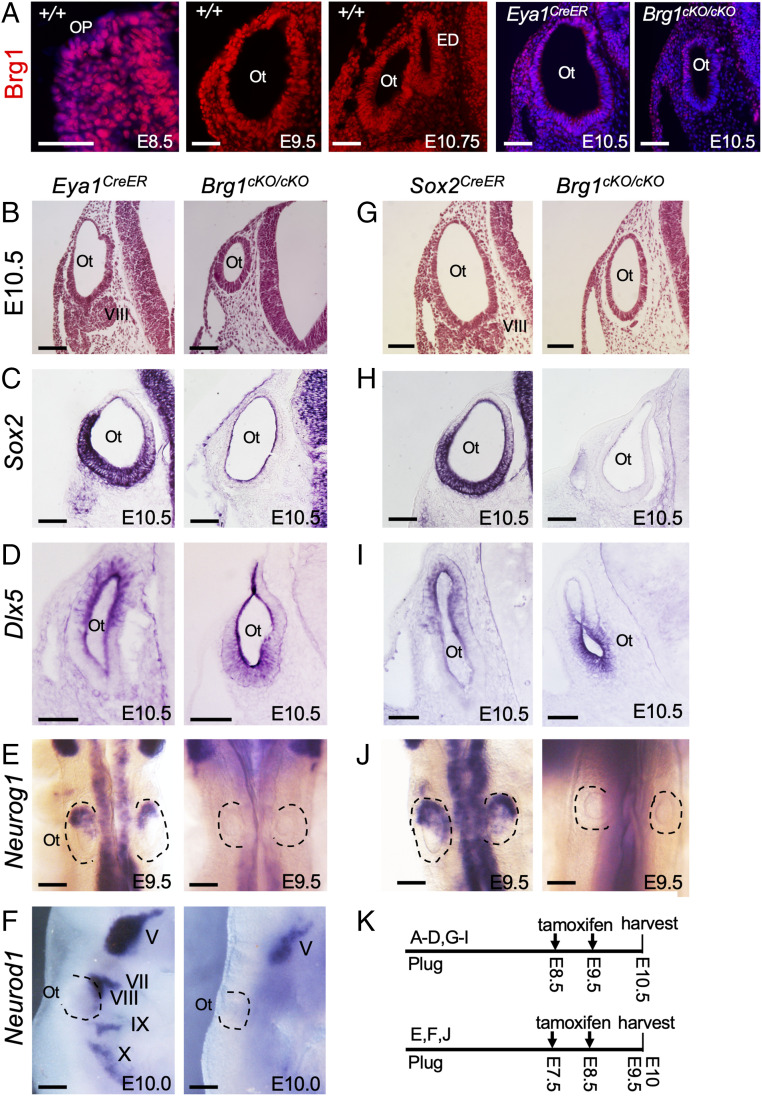
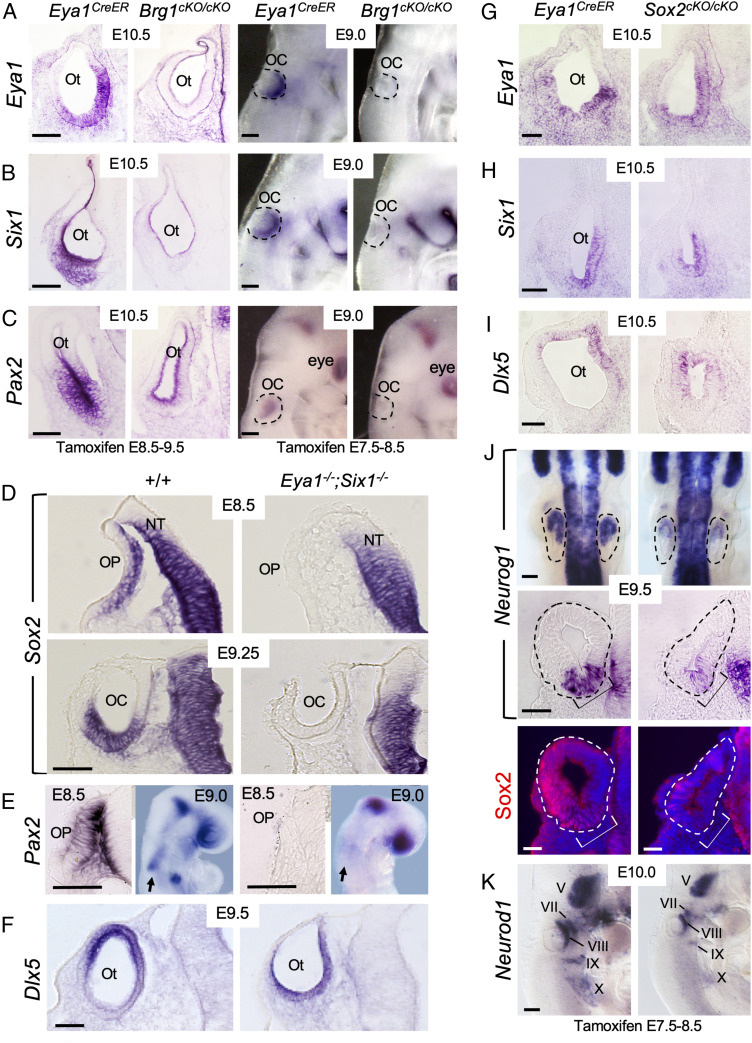
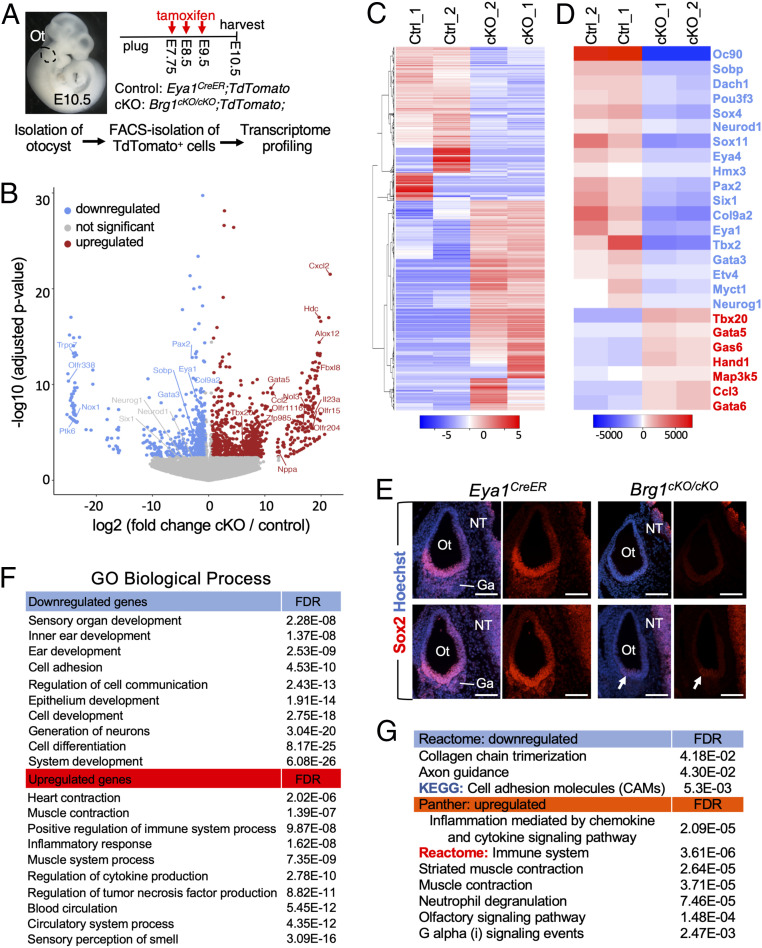
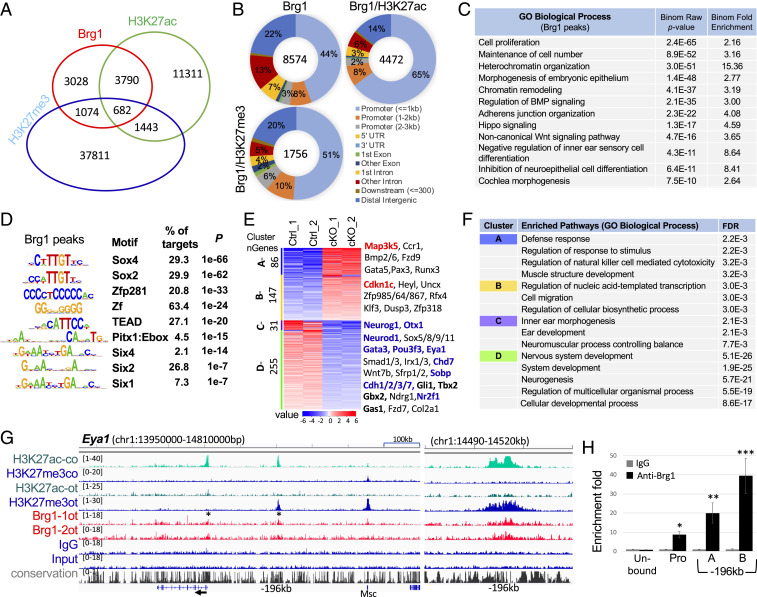
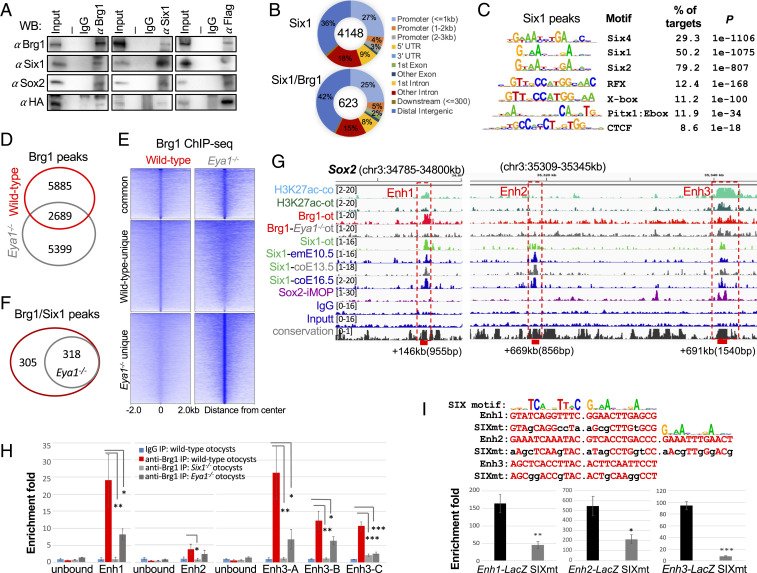
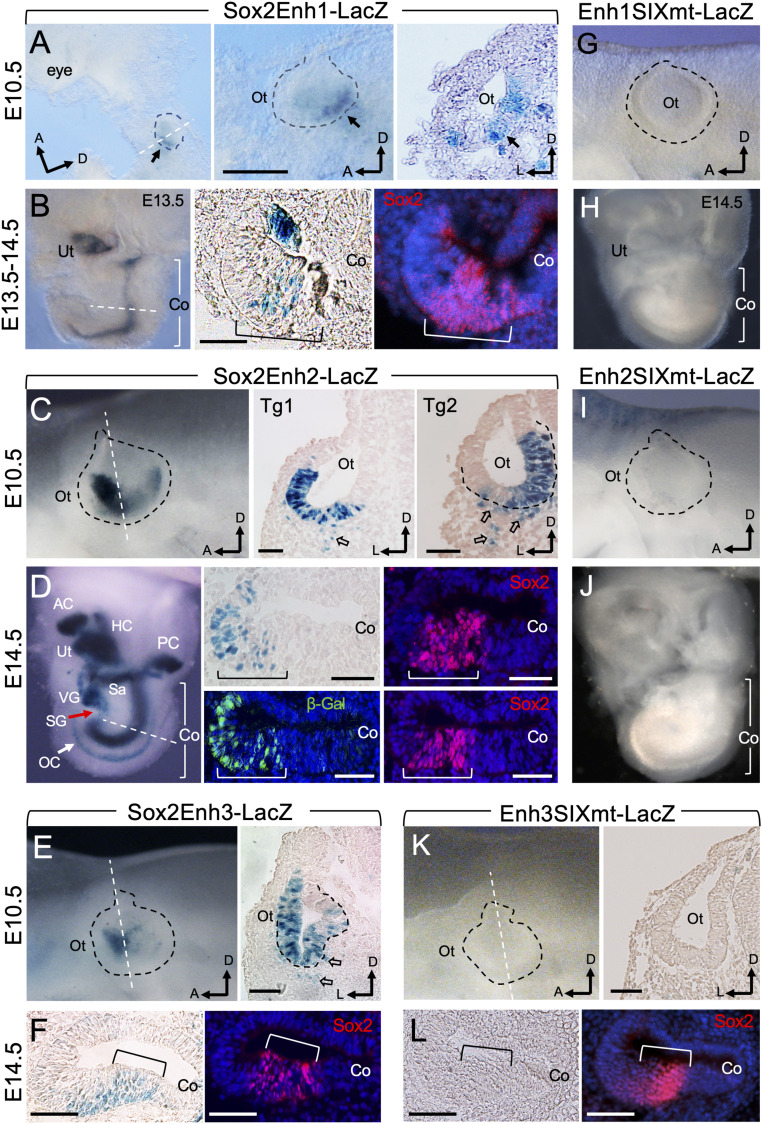
Specification of Sox2+ proneurosensory progenitors within otic ectoderm is a prerequisite for the production of sensory cells and neurons for hearing. However, the underlying molecular mechanisms driving this lineage specification remain unknown. Here, we show that the Brg1-based SWI/SNF chromatin-remodeling complex interacts with the neurosensory-specific transcriptional regulators Eya1/Six1 to induce Sox2 expression and promote proneurosensory-lineage specification. Ablation of the ATPase-subunit Brg1 or both Eya1/Six1 results in loss of Sox2 expression and lack of neurosensory identity, leading to abnormal apoptosis within the otic ectoderm. Brg1 binds to two of three distal 3' Sox2 enhancers occupied by Six1, and Brg1-binding to these regions depends on Eya1-Six1 activity. We demonstrate that the activity of these Sox2 enhancers in otic neurosensory cells specifically depends on binding to Six1. Furthermore, genome-wide and transcriptome profiling indicate that Brg1 may suppress apoptotic factor Map3k5 to inhibit apoptosis. Together, our findings reveal an essential role for Brg1, its downstream pathways, and their interactions with Six1/Eya1 in promoting proneurosensory fate induction in the otic ectoderm and subsequent neuronal lineage commitment and survival of otic cells.
Keywords: Sox2 enhancers; chromatin remodelers; otic neurosensory lineage; regulation of otic Sox2 expression; transcription factors.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Ma Q., Chen Z., del Barco Barrantes I., de la Pompa J. L., Anderson D. J., neurogenin1 is essential for the determination of neuronal precursors for proximal cranial sensory ganglia. Neuron 20, 469–482 (1998). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
