

The aryl hydrocarbon receptor facilitates the human cytomegalovirus-mediated G1/S block to cell cycle progression
- PMID: 33723080
- PMCID: PMC8000196
- DOI: 10.1073/pnas.2026336118
The aryl hydrocarbon receptor facilitates the human cytomegalovirus-mediated G1/S block to cell cycle progression
Abstract
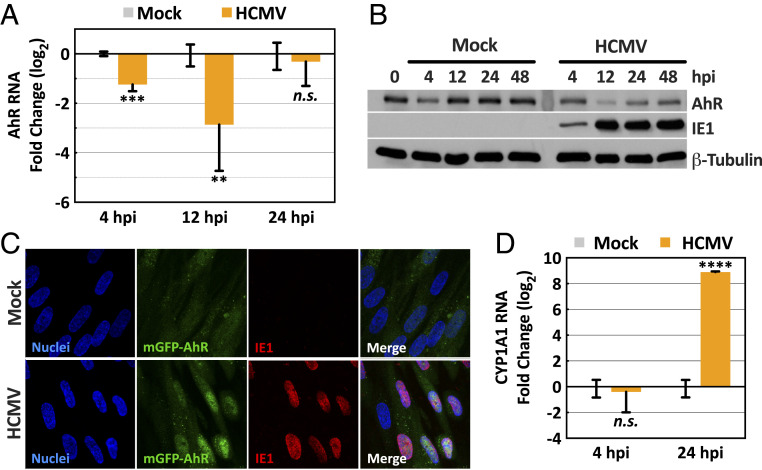
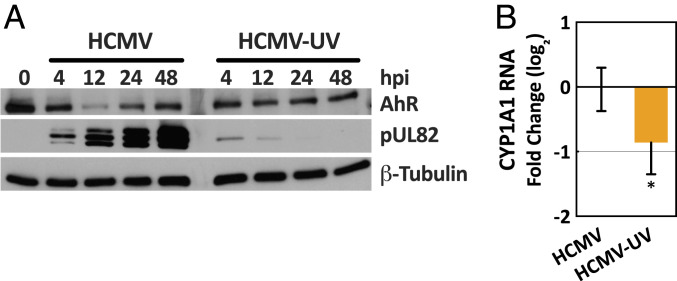
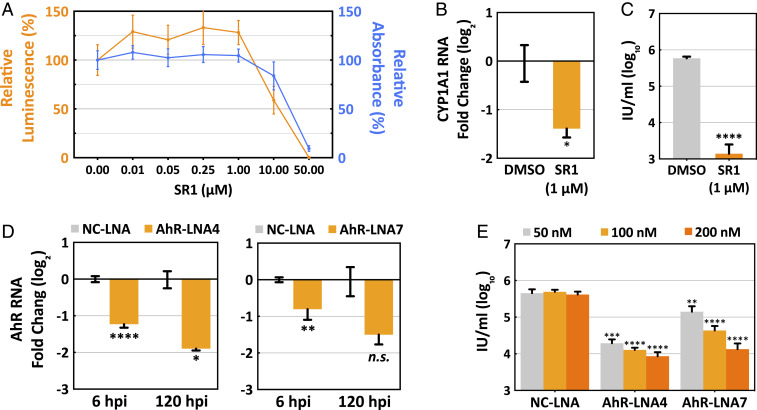
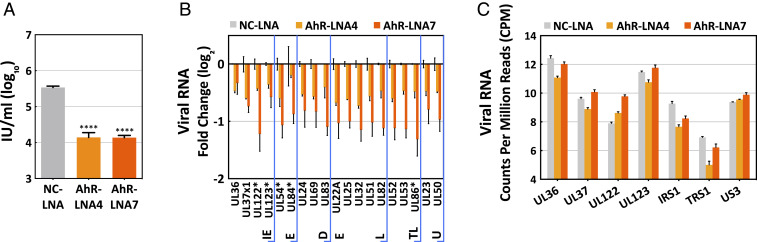
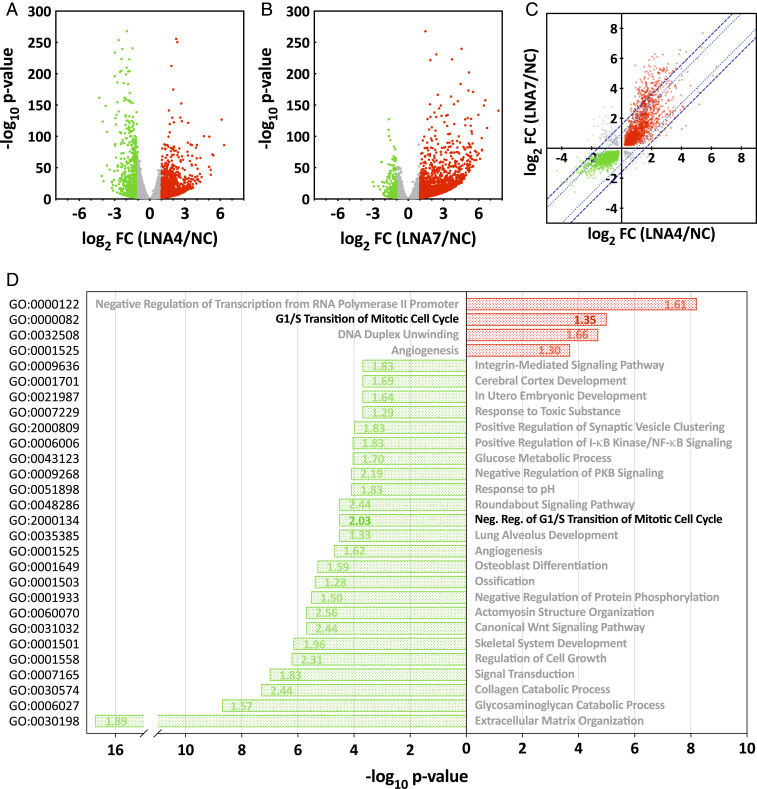
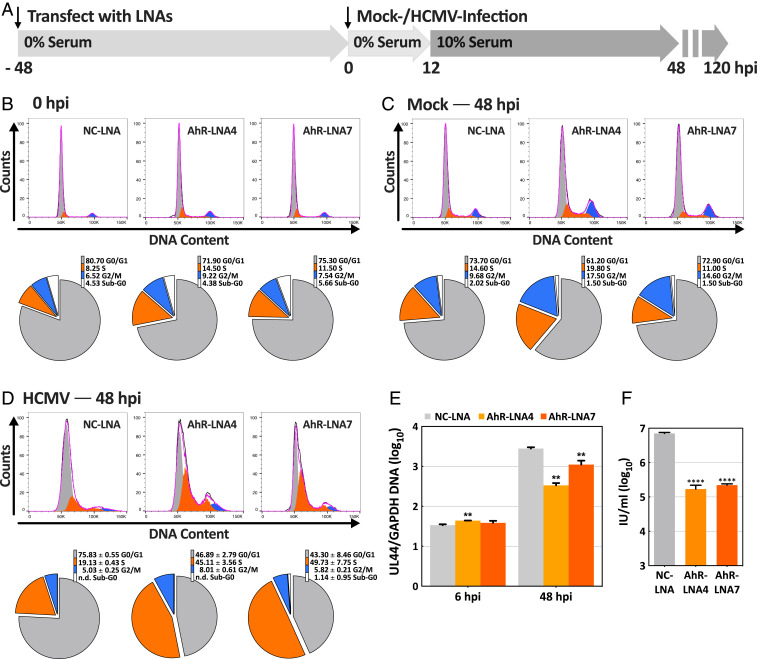
The tryptophan metabolite, kynurenine, is known to be produced at elevated levels within human cytomegalovirus (HCMV)-infected fibroblasts. Kynurenine is an endogenous aryl hydrocarbon receptor (AhR) ligand. Here we show that the AhR is activated following HCMV infection, and pharmacological inhibition of AhR or knockdown of AhR RNA reduced the accumulation of viral RNAs and infectious progeny. RNA-seq analysis of infected cells following AhR knockdown showed that the receptor alters the levels of numerous RNAs, including RNAs related to cell cycle progression. AhR knockdown alleviated the G1/S cell cycle block that is normally instituted in HCMV-infected fibroblasts, consistent with its known ability to regulate cell cycle progression and cell proliferation. In sum, AhR is activated by kynurenine and perhaps other ligands produced during HCMV infection, it profoundly alters the infected-cell transcriptome, and one outcome of its activity is a block to cell cycle progression, providing mechanistic insight to a long-known element of the virus-host cell interaction.
Keywords: aryl hydrocarbon receptor; cell cycle; human cytomegalovirus; kynurenine.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Griffiths P., Baraniak I., Reeves M., The pathogenesis of human cytomegalovirus. J. Pathol. 235, 288–297 (2015). - PubMed
-
- Cobbs C., Cytomegalovirus is a tumor-associated virus: Armed and dangerous. Curr. Opin. Virol. 39, 49–59 (2019). - PubMed
-
- Shenk T., Alwine J. C., Human cytomegalovirus: Coordinating cellular stress, signaling, and metabolic pathways. Annu. Rev. Virol. 1, 355–374 (2014). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
