

Single-cell atlas of early human brain development highlights heterogeneity of human neuroepithelial cells and early radial glia
- PMID: 33723434
- PMCID: PMC8012207
- DOI: 10.1038/s41593-020-00794-1
Single-cell atlas of early human brain development highlights heterogeneity of human neuroepithelial cells and early radial glia
Abstract
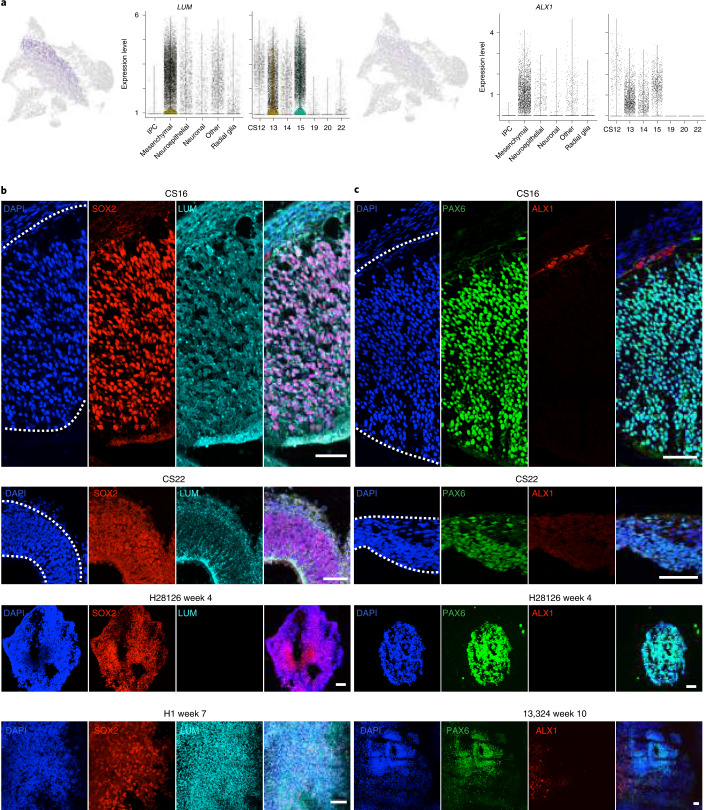
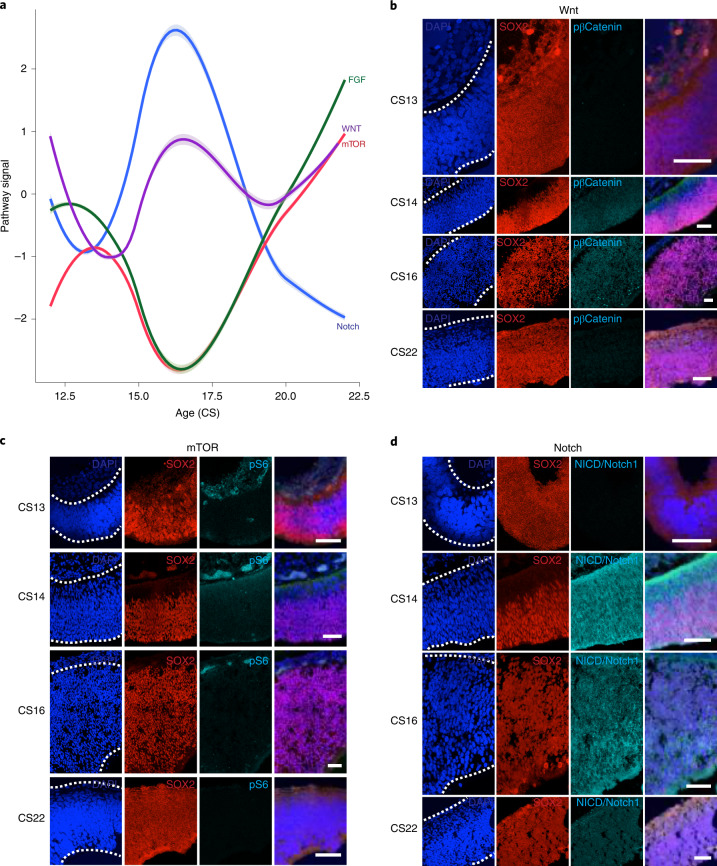
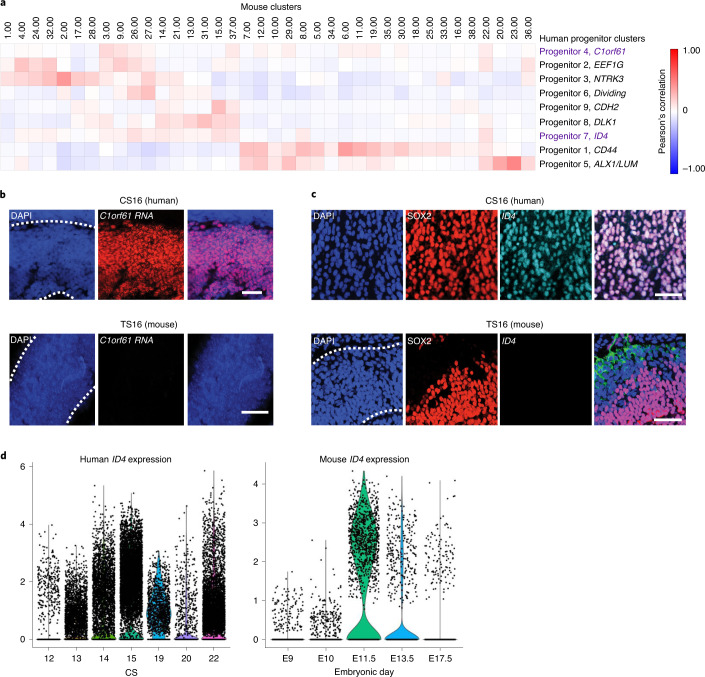
The human cortex comprises diverse cell types that emerge from an initially uniform neuroepithelium that gives rise to radial glia, the neural stem cells of the cortex. To characterize the earliest stages of human brain development, we performed single-cell RNA-sequencing across regions of the developing human brain, including the telencephalon, diencephalon, midbrain, hindbrain and cerebellum. We identify nine progenitor populations physically proximal to the telencephalon, suggesting more heterogeneity than previously described, including a highly prevalent mesenchymal-like population that disappears once neurogenesis begins. Comparison of human and mouse progenitor populations at corresponding stages identifies two progenitor clusters that are enriched in the early stages of human cortical development. We also find that organoid systems display low fidelity to neuroepithelial and early radial glia cell types, but improve as neurogenesis progresses. Overall, we provide a comprehensive molecular and spatial atlas of early stages of human brain and cortical development.
Conflict of interest statement
A.R.K. is a cofounder and board member of Neurona Therapeutics. The remaining authors declare no competing interests.
Figures


References
-
- Beccari L, Marco-Ferreres R, Bovolenta P. The logic of gene regulatory networks in early vertebrate forebrain patterning. Mech. Dev. 2013;130:95–111. - PubMed
-
- Gotz M, Hartfuss E, Malatesta P. Radial glial cells as neuronal precursors: a new perspective on the correlation of morphology and lineage restriction in the developing cerebral cortex of mice. Brain Res. Bull. 2002;57:777–788. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
