

A peroxisomal β-oxidative pathway contributes to the formation of C6-C1 aromatic volatiles in poplar
- PMID: 33723573
- PMCID: PMC8195509
- DOI: 10.1093/plphys/kiab111
A peroxisomal β-oxidative pathway contributes to the formation of C6-C1 aromatic volatiles in poplar
Abstract
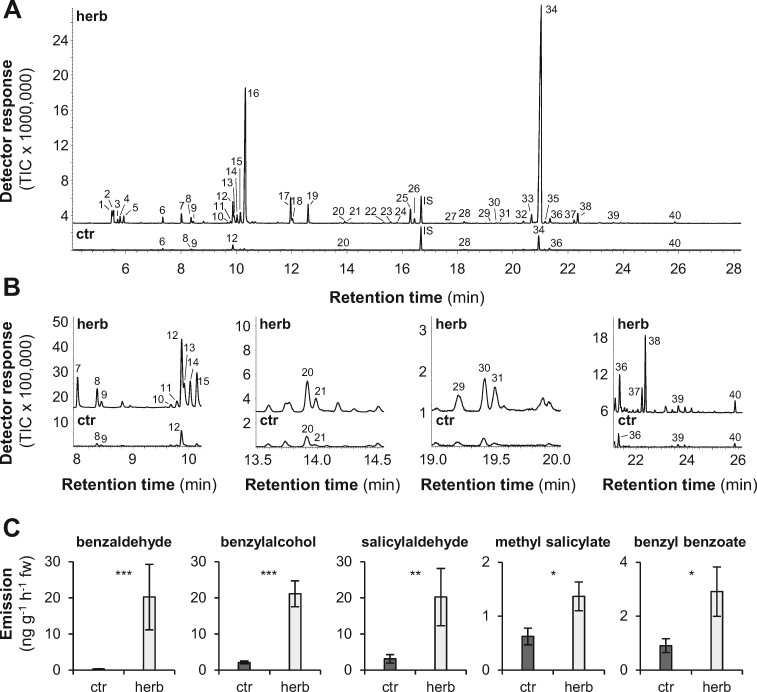
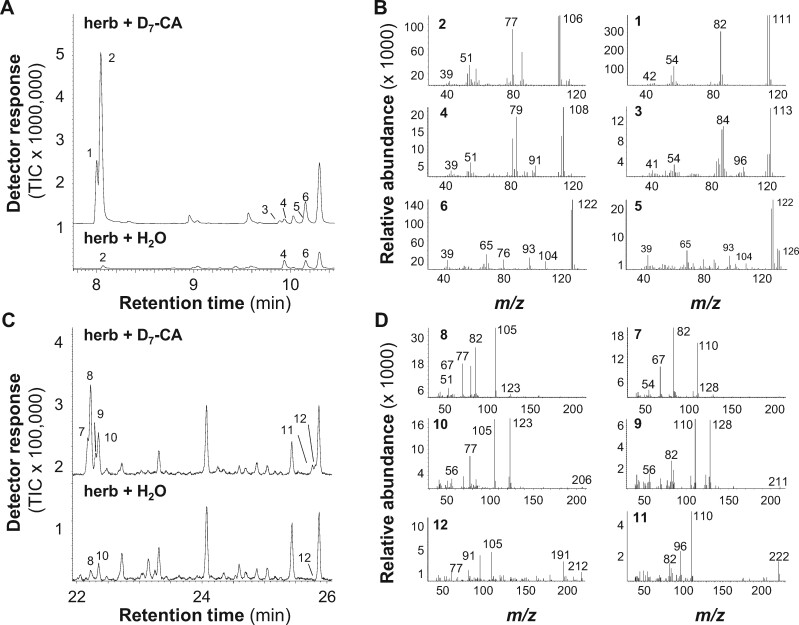
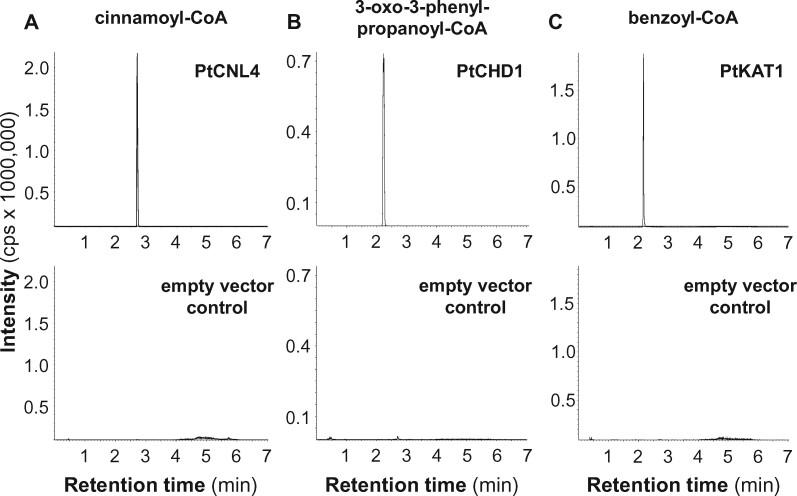
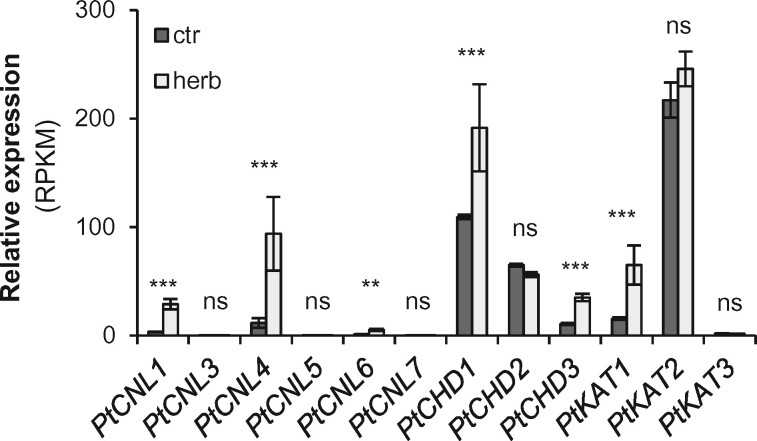
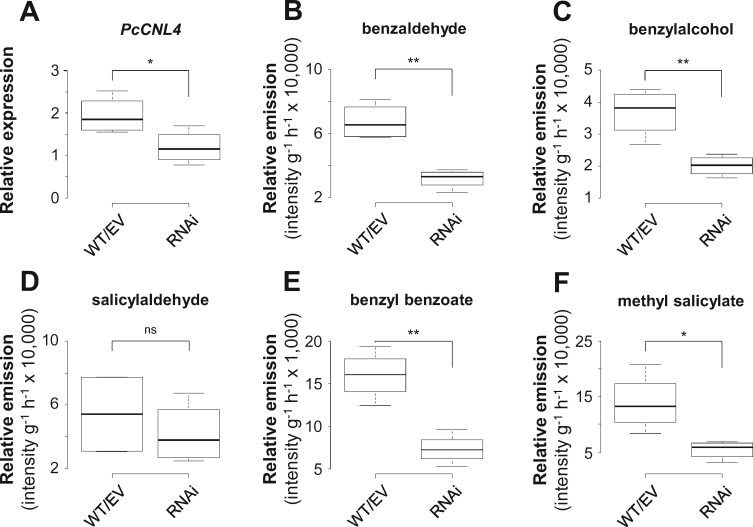
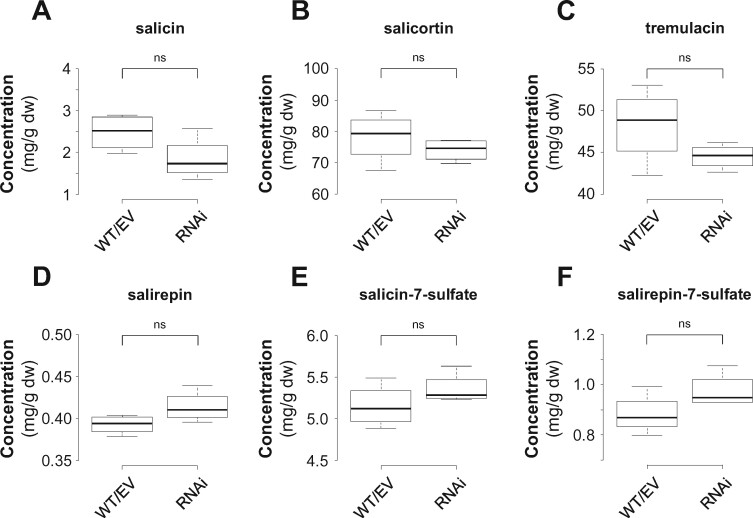
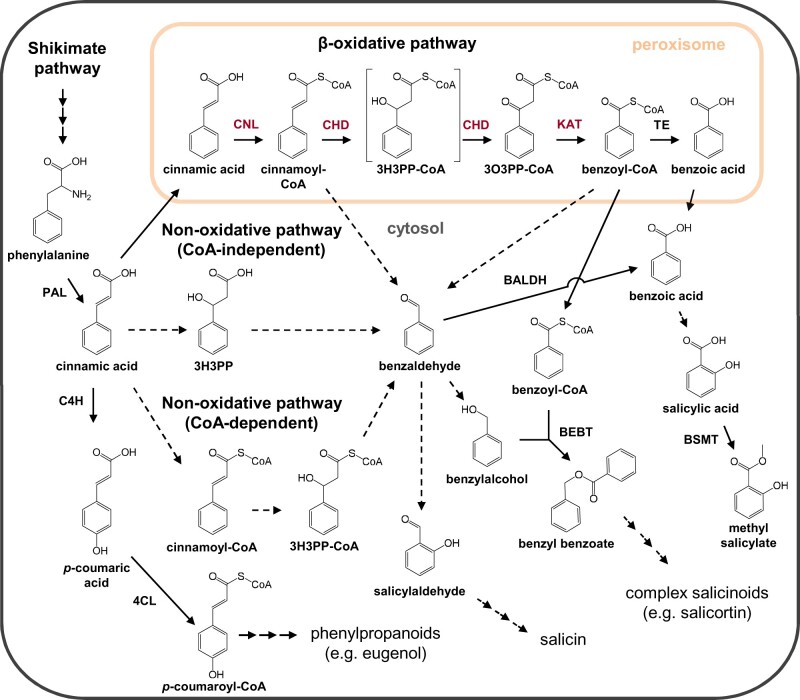
Benzenoids (C6-C1 aromatic compounds) play important roles in plant defense and are often produced upon herbivory. Black cottonwood (Populus trichocarpa) produces a variety of volatile and nonvolatile benzenoids involved in various defense responses. However, their biosynthesis in poplar is mainly unresolved. We showed feeding of the poplar leaf beetle (Chrysomela populi) on P. trichocarpa leaves led to increased emission of the benzenoid volatiles benzaldehyde, benzylalcohol, and benzyl benzoate. The accumulation of salicinoids, a group of nonvolatile phenolic defense glycosides composed in part of benzenoid units, was hardly affected by beetle herbivory. In planta labeling experiments revealed that volatile and nonvolatile poplar benzenoids are produced from cinnamic acid (C6-C3). The biosynthesis of C6-C1 aromatic compounds from cinnamic acid has been described in petunia (Petunia hybrida) flowers where the pathway includes a peroxisomal-localized chain shortening sequence, involving cinnamate-CoA ligase (CNL), cinnamoyl-CoA hydratase/dehydrogenase (CHD), and 3-ketoacyl-CoA thiolase (KAT). Sequence and phylogenetic analysis enabled the identification of small CNL, CHD, and KAT gene families in P. trichocarpa. Heterologous expression of the candidate genes in Escherichia coli and characterization of purified proteins in vitro revealed enzymatic activities similar to those described in petunia flowers. RNA interference-mediated knockdown of the CNL subfamily in gray poplar (Populus x canescens) resulted in decreased emission of C6-C1 aromatic volatiles upon herbivory, while constitutively accumulating salicinoids were not affected. This indicates the peroxisomal β-oxidative pathway participates in the formation of volatile benzenoids. The chain shortening steps for salicinoids, however, likely employ an alternative pathway.
© The Author(s) 2021. Published by Oxford University Press on behalf of American Society of Plant Biologists.
Figures
References
-
- Abd El-Mawla AM, Beerhues L (2002) Benzoic acid biosynthesis in cell cultures of Hypericum androsaemum. Planta 214: 727–733 - PubMed
-
- Adebesin F, Widhalm JR, Lynch JH, McCoy RM, Dudareva N (2018) A peroxisomal thioesterase plays auxiliary roles in plant β-oxidative benzoic acid metabolism. Plant J 93: 905–916 - PubMed
-
- Amrad A, Moser M, Mandel T, de Vries M, Schuurink RC, Freitas L, Kuhlemeier C (2016) Gain and loss of floral scent production through changes in structural genes during pollinator-mediated speciation. Curr Biol 26: 3303–3312 - PubMed
-
- Babst BA, Harding SA, Tsai C-J (2010) Biosynthesis of phenolic glycosides from phenylpropanoid and benzenoid precursors in Populus. J Chem Ecol 36: 286–297 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
