

Disruption of barley immunity to powdery mildew by an in-frame Lys-Leu deletion in the essential protein SGT1
- PMID: 33724411
- PMCID: PMC8045700
- DOI: 10.1093/genetics/iyaa026
Disruption of barley immunity to powdery mildew by an in-frame Lys-Leu deletion in the essential protein SGT1
Abstract
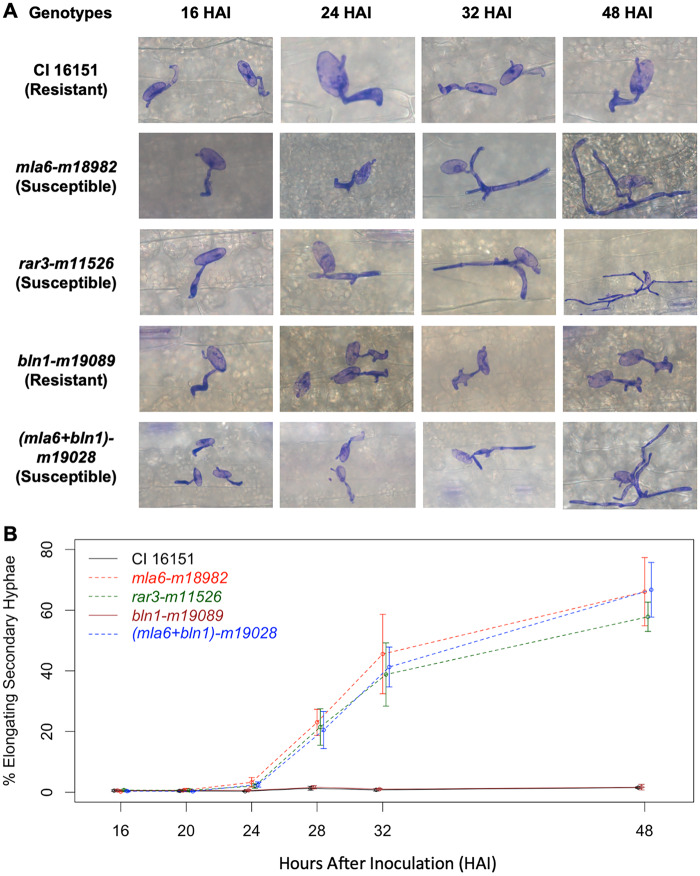
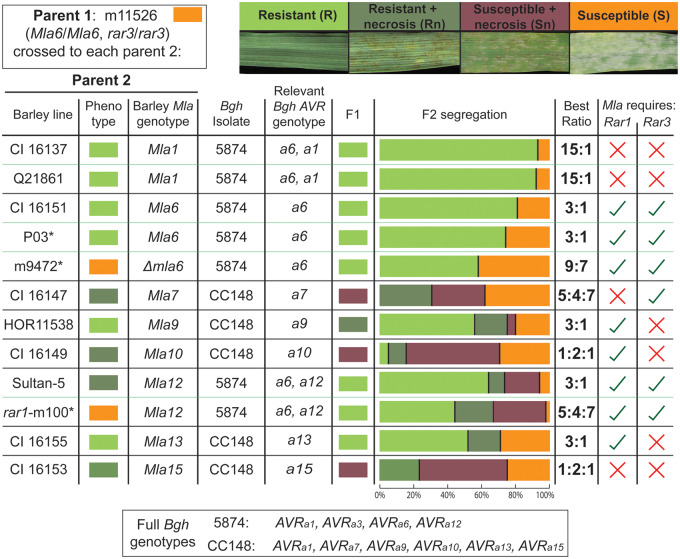
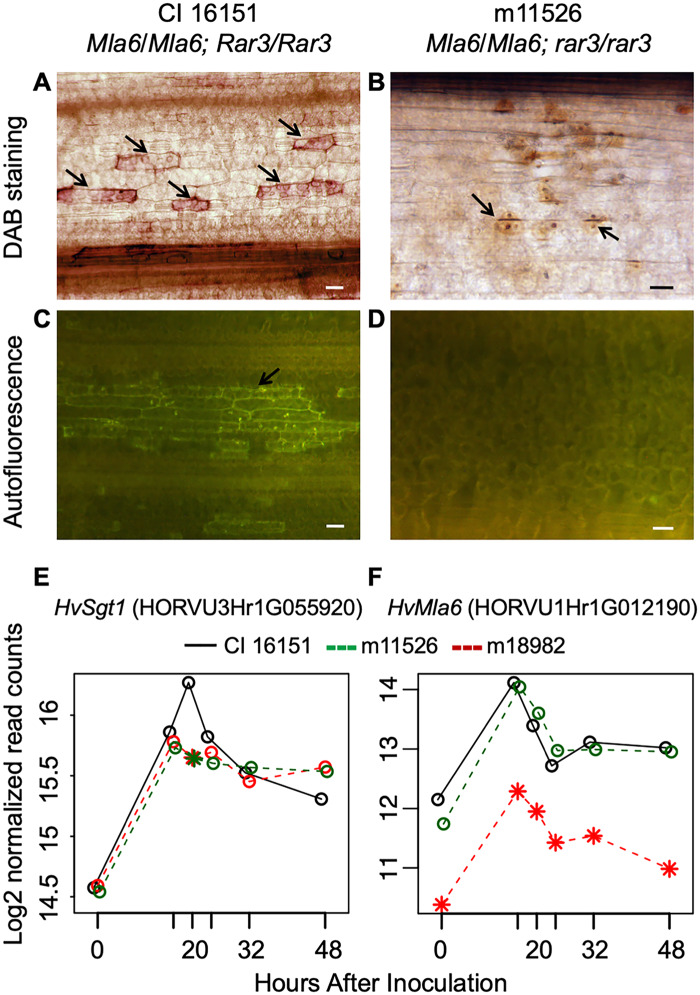
Barley (Hordeum vulgare L.) Mla (Mildew resistance locus a) and its nucleotide-binding, leucine-rich-repeat receptor (NLR) orthologs protect many cereal crops from diseases caused by fungal pathogens. However, large segments of the Mla pathway and its mechanisms remain unknown. To further characterize the molecular interactions required for NLR-based immunity, we used fast-neutron mutagenesis to screen for plants compromised in MLA-mediated response to the powdery mildew fungus, Blumeria graminis f. sp. hordei. One variant, m11526, contained a novel mutation, designated rar3 (required for Mla6 resistance3), that abolishes race-specific resistance conditioned by the Mla6, Mla7, and Mla12 alleles, but does not compromise immunity mediated by Mla1, Mla9, Mla10, and Mla13. This is analogous to, but unique from, the differential requirement of Mla alleles for the co-chaperone Rar1 (required for Mla12 resistance1). We used bulked-segregant-exome capture and fine mapping to delineate the causal mutation to an in-frame Lys-Leu deletion within the SGS domain of SGT1 (Suppressor of G-two allele of Skp1, Sgt1ΔKL308-309), the structural region that interacts with MLA proteins. In nature, mutations to Sgt1 usually cause lethal phenotypes, but here we pinpoint a unique modification that delineates its requirement for some disease resistances, while unaffecting others as well as normal cell processes. Moreover, the data indicate that the requirement of SGT1 for resistance signaling by NLRs can be delimited to single sites on the protein. Further study could distinguish the regions by which pathogen effectors and host proteins interact with SGT1, facilitating precise editing of effector incompatible variants.
Keywords: Blumeria graminis; SGT1; barley; in-frame deletion; leucine-rich-repeat receptor (NLR); nucleotide binding; resistance signaling.
Published by Oxford University Press on behalf of Genetics Society of America 2020. This work is written by US Government employees and is in the public domain in the US.
Figures


Similar articles
-
A single-amino acid substitution in the sixth leucine-rich repeat of barley MLA6 and MLA13 alleviates dependence on RAR1 for disease resistance signaling.Plant J. 2004 Apr;38(2):215-26. doi: 10.1111/j.1365-313X.2004.02032.x. Plant J. 2004. PMID: 15078326
-
SGT1-Specific Domain Mutations Impair Interactions with the Barley MLA6 Immune Receptor in Association with Loss of NLR Protein.Mol Plant Microbe Interact. 2022 Mar;35(3):274-289. doi: 10.1094/MPMI-08-21-0217-R. Epub 2022 Mar 18. Mol Plant Microbe Interact. 2022. PMID: 34889653
-
The leucine-rich repeats in allelic barley MLA immune receptors define specificity towards sequence-unrelated powdery mildew avirulence effectors with a predicted common RNase-like fold.PLoS Pathog. 2021 Feb 3;17(2):e1009223. doi: 10.1371/journal.ppat.1009223. eCollection 2021 Feb. PLoS Pathog. 2021. PMID: 33534797 Free PMC article.
-
Magical mystery tour: MLO proteins in plant immunity and beyond.New Phytol. 2014 Oct;204(2):273-81. doi: 10.1111/nph.12889. New Phytol. 2014. PMID: 25453131 Review.
-
Resistance genes in barley (Hordeum vulgare L.) and their identification with molecular markers.J Appl Genet. 2003;44(3):291-309. J Appl Genet. 2003. PMID: 12923305 Review.
Cited by
-
FINDER: an automated software package to annotate eukaryotic genes from RNA-Seq data and associated protein sequences.BMC Bioinformatics. 2021 Apr 20;22(1):205. doi: 10.1186/s12859-021-04120-9. BMC Bioinformatics. 2021. PMID: 33879057 Free PMC article.
-
Powdery mildew effectors AVRA1 and BEC1016 target the ER J-domain protein HvERdj3B required for immunity in barley.Mol Plant Pathol. 2024 May;25(5):e13463. doi: 10.1111/mpp.13463. Mol Plant Pathol. 2024. PMID: 38695677 Free PMC article.
-
The Brachypodium distachyon cold-acclimated plasma membrane proteome is primed for stress resistance.G3 (Bethesda). 2021 Sep 6;11(9):jkab198. doi: 10.1093/g3journal/jkab198. G3 (Bethesda). 2021. PMID: 34544140 Free PMC article.
-
Characterization of the Roles of SGT1/RAR1, EDS1/NDR1, NPR1, and NRC/ADR1/NRG1 in Sw-5b-Mediated Resistance to Tomato Spotted Wilt Virus.Viruses. 2021 Jul 25;13(8):1447. doi: 10.3390/v13081447. Viruses. 2021. PMID: 34452313 Free PMC article.
-
An interolog-based barley interactome as an integration framework for immune signaling.Genetics. 2022 May 31;221(2):iyac056. doi: 10.1093/genetics/iyac056. Genetics. 2022. PMID: 35435213 Free PMC article.
References
-
- Aartsma-Rus A, Van Deutekom JCT, Fokkema IF, Van Ommen G-JB, Den Dunnen JT. 2006. Entries in the Leiden Duchenne muscular dystrophy mutation database: an overview of mutation types and paradoxical cases that confirm the reading-frame rule. Muscle Nerve. 34:135–144. - PubMed
-
- Austin MJ, Muskett P, Kahn K, Feys BJ, Jones JDG, et al. 2002. Regulatory role of SGT1 in early R gene-mediated plant defenses. Science. 295:2077–2080. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
