

A fish with no sex: gonadal and adrenal functions partition between zebrafish NR5A1 co-orthologs
- PMID: 33724412
- PMCID: PMC8045690
- DOI: 10.1093/genetics/iyaa030
A fish with no sex: gonadal and adrenal functions partition between zebrafish NR5A1 co-orthologs
Erratum in
-
Erratum to: A fish with no sex: gonadal and adrenal functions partition between zebrafish NR5A1 co-orthologs.Genetics. 2021 May 17;218(1):iyab051. doi: 10.1093/genetics/iyab051. Genetics. 2021. PMID: 33826717 Free PMC article. No abstract available.
Abstract
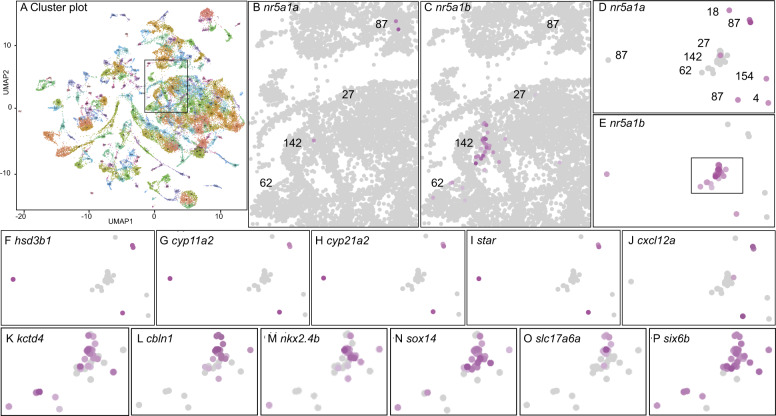
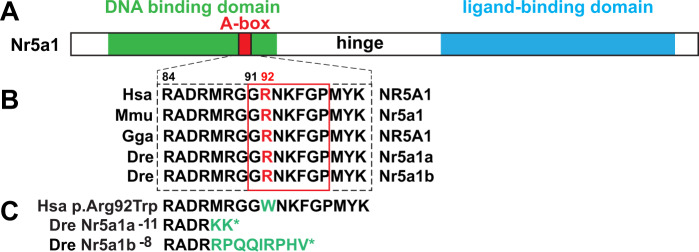
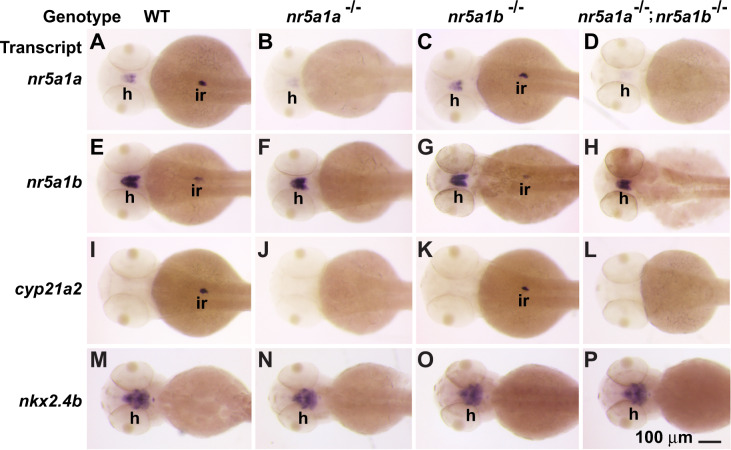
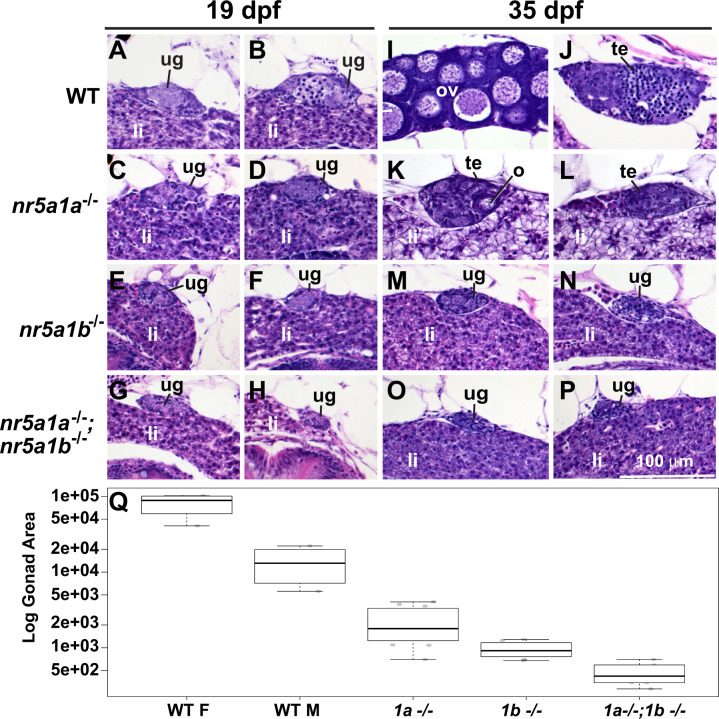
People with NR5A1 mutations experience testicular dysgenesis, ovotestes, or adrenal insufficiency, but we do not completely understand the origin of this phenotypic diversity. NR5A1 is expressed in gonadal soma precursor cells before expression of the sex-determining gene SRY. Many fish have two co-orthologs of NR5A1 that likely partitioned ancestral gene subfunctions between them. To explore ancestral roles of NR5A1, we knocked out nr5a1a and nr5a1b in zebrafish. Single-cell RNA-seq identified nr5a1a-expressing cells that co-expressed genes for steroid biosynthesis and the chemokine receptor Cxcl12a in 1-day postfertilization (dpf) embryos, as does the mammalian adrenal-gonadal (interrenal-gonadal) primordium. In 2dpf embryos, nr5a1a was expressed stronger in the interrenal-gonadal primordium than in the early hypothalamus but nr5a1b showed the reverse. Adult Leydig cells expressed both ohnologs and granulosa cells expressed nr5a1a stronger than nr5a1b. Mutants for nr5a1a lacked the interrenal, formed incompletely differentiated testes, had no Leydig cells, and grew far larger than normal fish. Mutants for nr5a1b formed a disorganized interrenal and their gonads completely disappeared. All homozygous mutant genotypes lacked secondary sex characteristics, including male breeding tubercles and female sex papillae, and had exceedingly low levels of estradiol, 11-ketotestosterone, and cortisol. RNA-seq showed that at 21dpf, some animals were developing as females and others were not, independent of nr5a1 genotype. By 35dpf, all mutant genotypes greatly under-expressed ovary-biased genes. Because adult nr5a1a mutants form gonads but lack an interrenal and conversely, adult nr5a1b mutants lack a gonad but have an interrenal, the adrenal, and gonadal functions of the ancestral nr5a1 gene partitioned between ohnologs after the teleost genome duplication, likely owing to reciprocal loss of ancestral tissue-specific regulatory elements. Identifying such elements could provide hints to otherwise unexplained cases of Differences in Sex Development.
Keywords: Differences in Sex Development; Genetics of Sex; SF1; adreno-gonadal primordium; disorders of sex development; scRNA-seq; subfunctionalization.
© The Author(s) 2020. Published by Oxford University Press on behalf of Genetics Society of America. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Figures

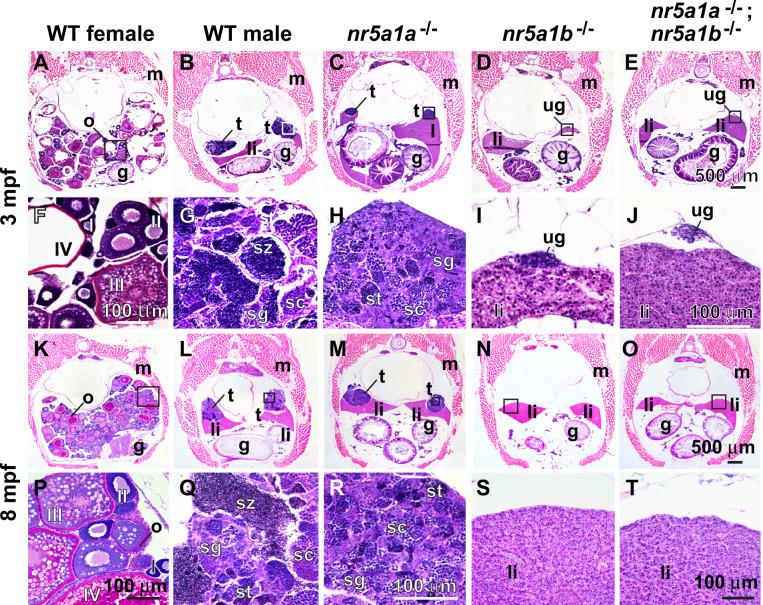
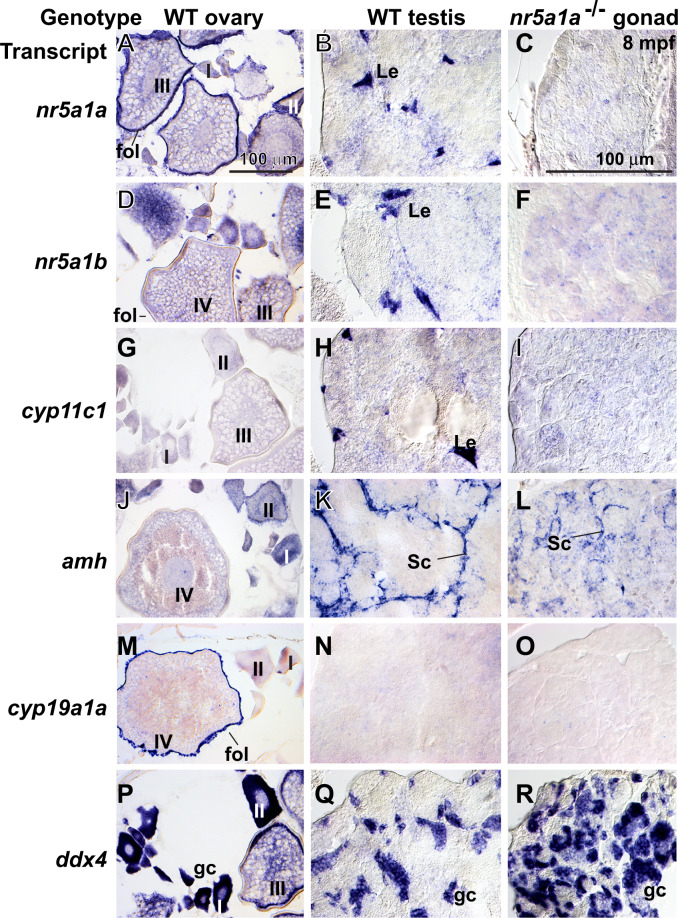
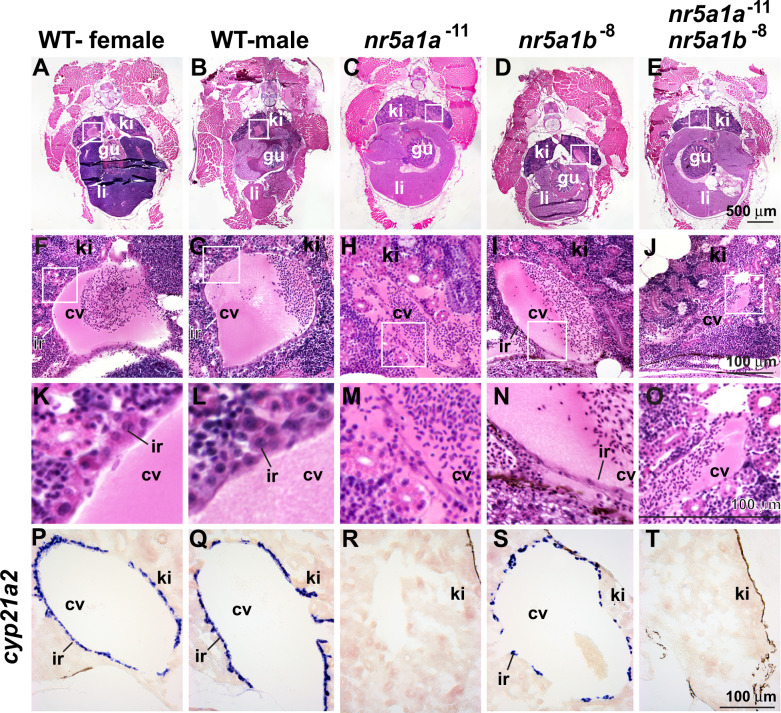
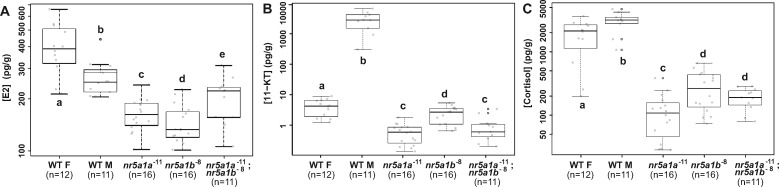
References
-
- Achermann JC, Ito M, Ito M, Hindmarsh PC, Jameson JL. 1999. A mutation in the gene encoding steroidogenic factor-1 causes XY sex reversal and adrenal failure in humans. Nat Genet. 22:125–126. - PubMed
-
- Amores A, Force A, Yan YL, Joly L, Amemiya C, et al. 1998. Zebrafish hox clusters and vertebrate genome evolution. Science. 282:1711–1714. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
