

Effects of targeting the transcription factors Ikaros and Aiolos on B cell activation and differentiation in systemic lupus erythematosus
- PMID: 33727237
- PMCID: PMC7970264
- DOI: 10.1136/lupus-2020-000445
Effects of targeting the transcription factors Ikaros and Aiolos on B cell activation and differentiation in systemic lupus erythematosus
Abstract
Objective: To evaluate the effects of targeting Ikaros and Aiolos by cereblon modulator iberdomide on the activation and differentiation of B-cells from patients with systemic lupus erythematosus (SLE).
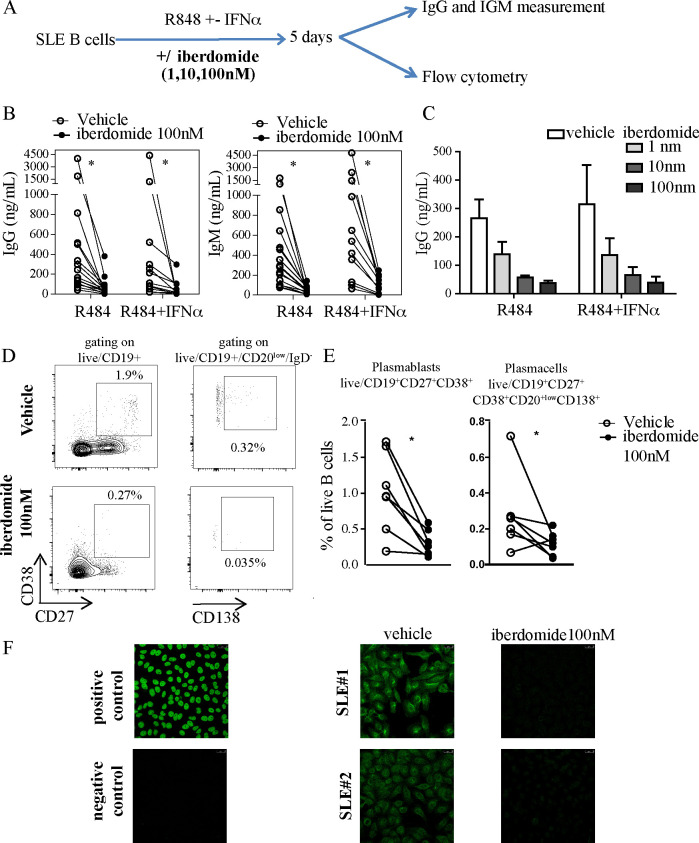
Methods: CD19+ B-cells isolated from the peripheral blood of patients with SLE (n=41) were cultured with TLR7 ligand resiquimod ±IFNα together with iberdomide or control from day 0 (n=16). Additionally, in vitro B-cell differentiation was induced by stimulation with IL-2/IL-10/IL-15/CD40L/resiquimod with iberdomide or control, given at day 0 or at day 4. At day 5, immunoglobulins were measured by ELISA and cells analysed by flow cytometry. RNA-Seq was performed on fluorescence-activated cell-sorted CD27-IgD+ naïve-B-cells and CD20lowCD27+CD38+ plasmablasts to investigate the transcriptional consequences of iberdomide.
Results: Iberdomide significantly inhibited the TLR7 and IFNα-mediated production of immunoglobulins from SLE B-cells and the production of antinuclear antibodies as well as significantly reducing the number of CD27+CD38+ plasmablasts (0.3±0.18, vehicle 1.01±0.56, p=0.011) and CD138+ plasma cells (0.12±0.06, vehicle 0.28±0.02, p=0.03). Additionally, treatment with iberdomide from day 0 significantly inhibited the differentiation of SLE B-cells into plasmablasts (6.4±13.5 vs vehicle 34.9±20.1, p=0.013) and antibody production. When given at later stages of differentiation, iberdomide did not affect the numbers of plasmablasts or the production of antibodies; however, it induced a significant modulation of gene expression involving IKZF1 and IKZF3 transcriptional programmes in both naïve B-cells and plasmablasts (400 and 461 differentially modulated genes, respectively, false discovery rate<0.05).
Conclusion: These results demonstrate the relevance of Ikaros and Aiolos as therapeutic targets in SLE due to their ability to modulate B cell activation and differentiation downstream of TLR7.
Keywords: B-lymphocytes; autoimmune diseases; lupus erythematosus; systemic.
© Author(s) (or their employer(s)) 2021. Re-use permitted under CC BY. Published by BMJ.
Conflict of interest statement
Competing interests: FR, MB, CP and ML received unrestricted grant support from Celgene. PS is an employee and shareholder of Celgene Corporation, now part of Bristol Myers Squibb.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials