

The role of SAMM50 in non-alcoholic fatty liver disease: from genetics to mechanisms
- PMID: 33728819
- PMCID: PMC8255833
- DOI: 10.1002/2211-5463.13146
The role of SAMM50 in non-alcoholic fatty liver disease: from genetics to mechanisms
Abstract
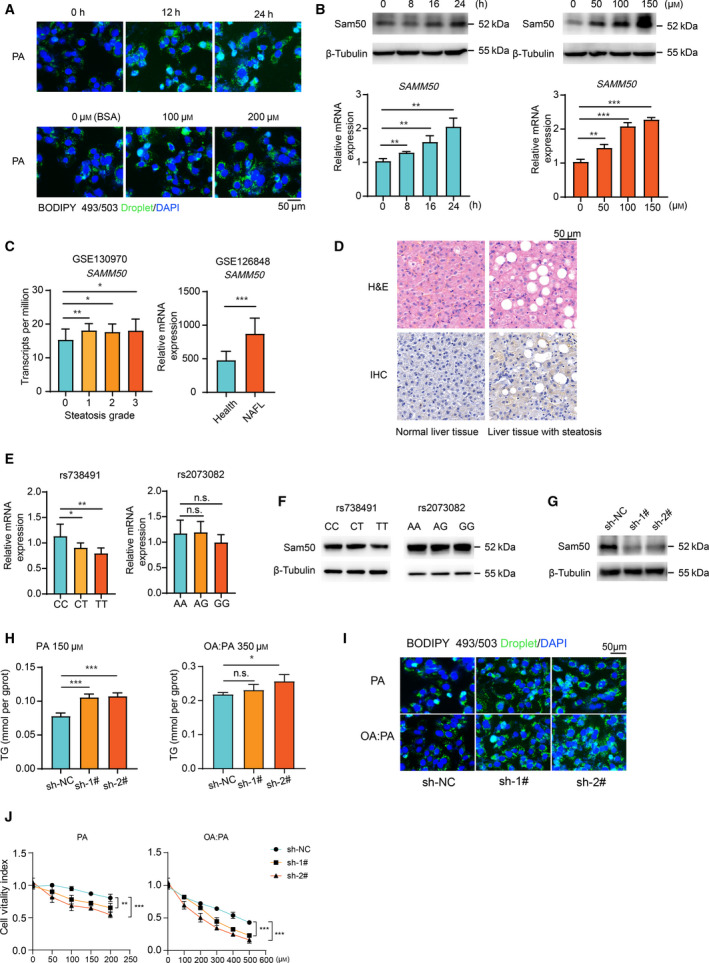
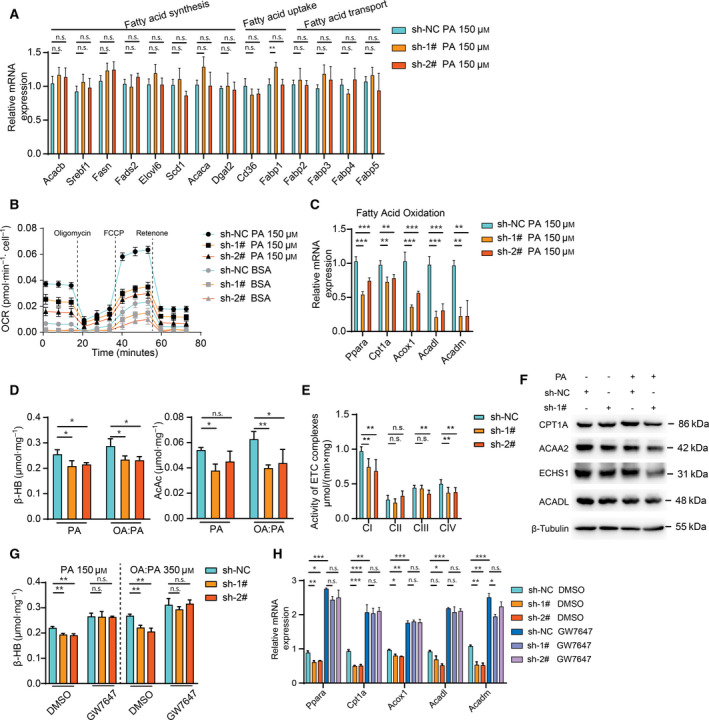
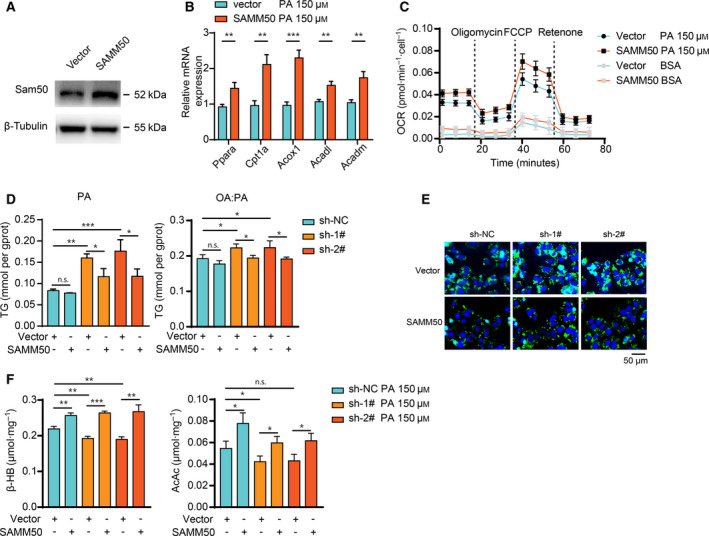
Non-alcoholic fatty liver disease (NAFLD) is characterized by hepatic lipid accumulation. SAMM50 encodes Sam50, a mitochondrial outer membrane protein involved in the removal of reactive oxygen species, mitochondrial morphology and regulation of mitophagy. Certain single nucleotide polymorphisms of SAMM50 have been reported to be correlated with NAFLD. However, the contribution of SAMM50 polymorphisms to the occurrence and severity of fatty liver in the Chinese Han cohort has rarely been reported. Here, we investigated the association between SAMM50 polymorphisms (rs738491 and rs2073082) and NAFLD in a Chinese Han cohort, as well as the mechanistic basis of this association. Clinical information and blood samples were collected from 380 NAFLD cases and 380 normal subjects for the detection of genotypes and biochemical parameters. Carriers of the rs738491 T allele or rs2073082 G allele of SAMM50 exhibit increased susceptibility to NAFLD [odds ratio (OR) = 1.39; 95% confidence interval (CI) = 1.14-1.71, P = 0.001; OR = 1.31; 95% CI = 1.05-1.62, P = 0.016, respectively] and are correlated with elevated serum triglyceride, alanine aminotransferase and aspartate aminotransferase levels. The presence of the T allele (TT + CT) of rs738491 (P < 0.01) or G allele (AG + GG) of rs2073082 (P = 0.03) is correlated with the severity of fatty liver in the NAFLD cohort. In vitro studies indicated that SAMM50 gene polymorphisms decrease its expression and SAMM50 deficiency results in increased lipid accumulation as a result of a decrease in fatty acid oxidation. Overexpression of SAMM50 enhances fatty acid oxidation and mitigates intracellular lipid accumulation. Our results confirm the association between the SAMM50 rs738491 and rs2073082 polymorphisms and the risk of fatty liver in a Chinese cohort. The underlying mechanism may be related to decreased fatty acid oxidation caused by SAMM50 deficiency.
Keywords: SAMM50; NAFLD; SNPs; fatty acid oxidation.
© 2021 The Authors. FEBS Open Bio published by John Wiley & Sons Ltd on behalf of Federation of European Biochemical Societies.
Conflict of interest statement
The authors declare that they have no conflicts of interest.
Figures
Similar articles
-
SAMM50-rs2073082, -rs738491 and -rs3761472 Interactions Enhancement of Susceptibility to Non-Alcoholic Fatty Liver Disease.Biomedicines. 2023 Aug 29;11(9):2416. doi: 10.3390/biomedicines11092416. Biomedicines. 2023. PMID: 37760857 Free PMC article.
-
Genetic Variants in the SAMM50 Gene Create Susceptibility to Nonalcoholic Fatty Liver Disease in a Chinese Han Population.Hepat Mon. 2015 Oct 10;15(10):e31076. doi: 10.5812/hepatmon.31076. eCollection 2015 Oct. Hepat Mon. 2015. PMID: 26587038 Free PMC article.
-
Genome-wide scan revealed that polymorphisms in the PNPLA3, SAMM50, and PARVB genes are associated with development and progression of nonalcoholic fatty liver disease in Japan.Hum Genet. 2013 Jul;132(7):783-92. doi: 10.1007/s00439-013-1294-3. Epub 2013 Mar 28. Hum Genet. 2013. PMID: 23535911 Clinical Trial.
-
Genetics of nonalcoholic fatty liver disease in Asian populations.J Genet. 2019 Mar;98:29. J Genet. 2019. PMID: 30945694 Review.
-
Update on Non-Alcoholic Fatty Liver Disease-Associated Single Nucleotide Polymorphisms and Their Involvement in Liver Steatosis, Inflammation, and Fibrosis: A Narrative Review.Iran Biomed J. 2022 Jul 1;26(4):252-68. doi: 10.52547/ibj.3647. Iran Biomed J. 2022. PMID: 36000237 Free PMC article. Review.
Cited by
-
Vitamin D ameliorates age-induced nonalcoholic fatty liver disease by increasing the mitochondrial contact site and cristae organizing system (MICOS) 60 level.Exp Mol Med. 2024 Feb;56(1):142-155. doi: 10.1038/s12276-023-01125-7. Epub 2024 Jan 4. Exp Mol Med. 2024. PMID: 38172593 Free PMC article.
-
Ablation of Sam50 is associated with fragmentation and alterations in metabolism in murine and human myotubes.bioRxiv [Preprint]. 2023 Oct 19:2023.05.20.541602. doi: 10.1101/2023.05.20.541602. bioRxiv. 2023. PMID: 37292887 Free PMC article. Preprint.
-
Mitochondrial DNA release and sensing in innate immune responses.Hum Mol Genet. 2024 May 22;33(R1):R80-R91. doi: 10.1093/hmg/ddae031. Hum Mol Genet. 2024. PMID: 38779772 Free PMC article. Review.
-
Newly identified single-nucleotide polymorphism associated with the transition from nonalcoholic fatty liver disease to liver fibrosis: results from a nested case-control study in the UK biobank.Ann Med. 2025 Dec;57(1):2458201. doi: 10.1080/07853890.2025.2458201. Epub 2025 Feb 3. Ann Med. 2025. PMID: 39898988 Free PMC article.
-
SAMM50-rs2073082, -rs738491 and -rs3761472 Interactions Enhancement of Susceptibility to Non-Alcoholic Fatty Liver Disease.Biomedicines. 2023 Aug 29;11(9):2416. doi: 10.3390/biomedicines11092416. Biomedicines. 2023. PMID: 37760857 Free PMC article.
References
-
- Trepo E and Valenti L (2020) Update on NAFLD genetics: from new variants to the clinic. J Hepatol 72, 1196–1209. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
