

Mesenchymal stem cell-derived extracellular vesicles reduce senescence and extend health span in mouse models of aging
- PMID: 33728821
- PMCID: PMC8045949
- DOI: 10.1111/acel.13337
Mesenchymal stem cell-derived extracellular vesicles reduce senescence and extend health span in mouse models of aging
Abstract
Aging drives progressive loss of the ability of tissues to recover from stress, partly through loss of somatic stem cell function and increased senescent burden. We demonstrate that bone marrow-derived mesenchymal stem cells (BM-MSCs) rapidly senescence and become dysfunctional in culture. Injection of BM-MSCs from young mice prolonged life span and health span, and conditioned media (CM) from young BM-MSCs rescued the function of aged stem cells and senescent fibroblasts. Extracellular vesicles (EVs) from young BM-MSC CM extended life span of Ercc1-/- mice similarly to injection of young BM-MSCs. Finally, treatment with EVs from MSCs generated from human ES cells reduced senescence in culture and in vivo, and improved health span. Thus, MSC EVs represent an effective and safe approach for conferring the therapeutic effects of adult stem cells, avoiding the risks of tumor development and donor cell rejection. These results demonstrate that MSC-derived EVs are highly effective senotherapeutics, slowing the progression of aging, and diseases driven by cellular senescence.
Keywords: aging; extracellular vesicles; mesenchymal stem cells; senescence; stem cells.
© 2021 The Authors. Aging Cell published by the Anatomical Society and John Wiley & Sons Ltd.
Conflict of interest statement
SKL is a founder of Paracrine Therapeutics, which develops EVs for therapeutic applications. PDR and LJN are co‐founders of NRTK Biosciences, which is developing approaches to reduce the senescent cell burden.
Figures
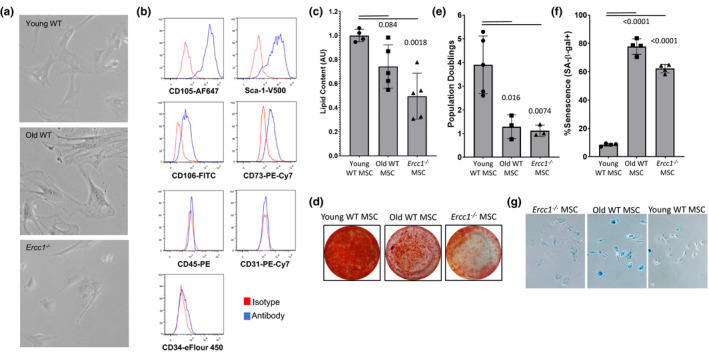
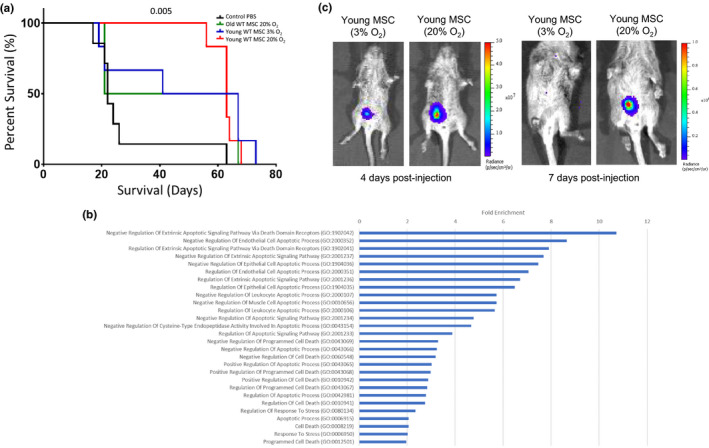
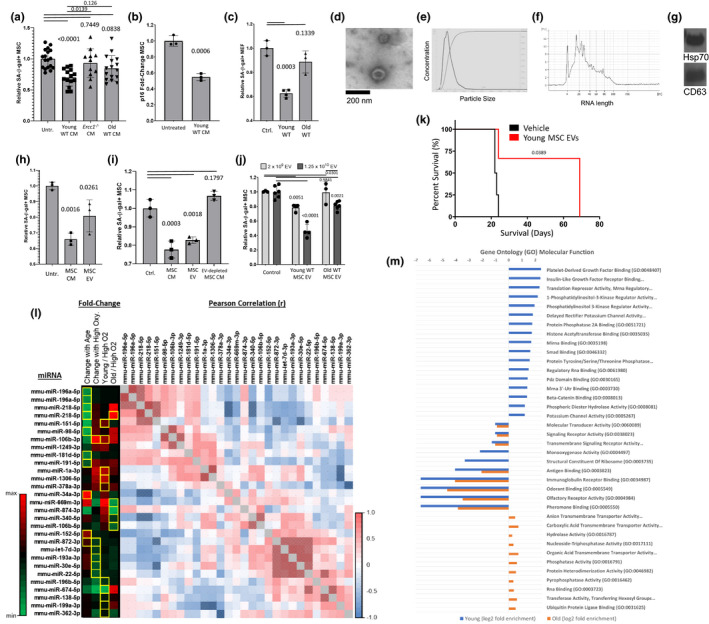
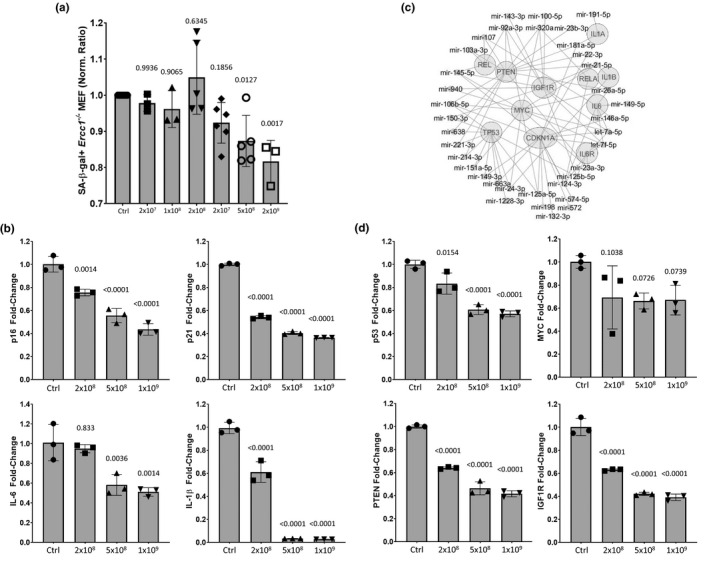
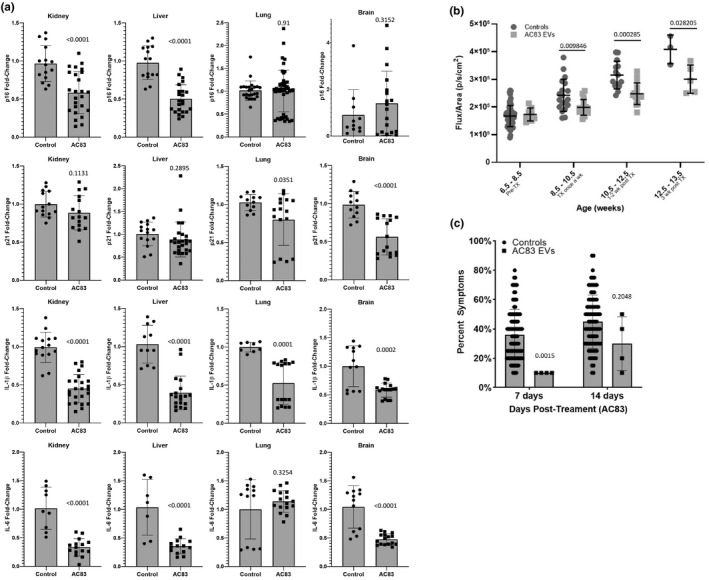
 ) correlated. (m) Gene ontology (GO; molecular function) of miRNA targetomes for miRNAs enriched in young and old MSC EVs; axis is log2 of fold enrichment and all categories are significant (p < 0.05) after false‐discovery rate (FDR) correction. Raw miRNA read counts, pairwise Pearson r and p values, and confidence intervals available in source data file
) correlated. (m) Gene ontology (GO; molecular function) of miRNA targetomes for miRNAs enriched in young and old MSC EVs; axis is log2 of fold enrichment and all categories are significant (p < 0.05) after false‐discovery rate (FDR) correction. Raw miRNA read counts, pairwise Pearson r and p values, and confidence intervals available in source data file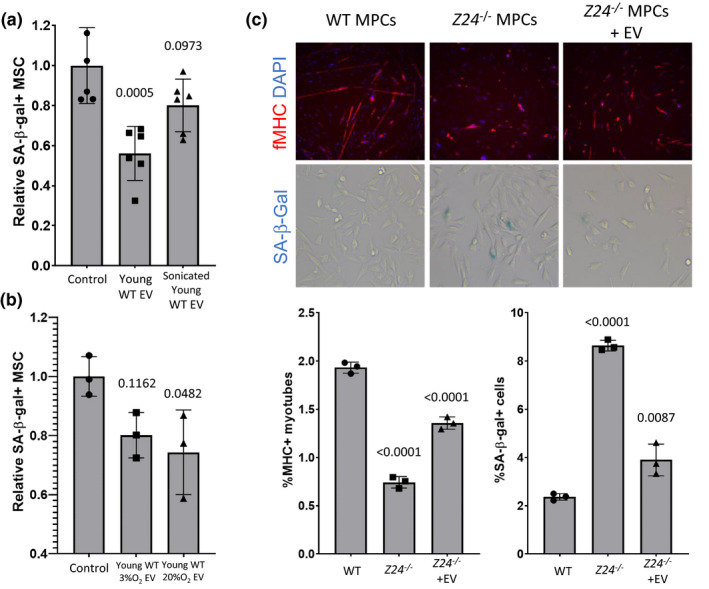
References
-
- Arslan, F. , Lai, R. C. , Smeets, M. B. , Akeroyd, L. , Choo, A. , Aguor, E. N. , Timmers, L. , van Rijen, H. V. , Doevendans, P. A. , Pasterkamp, G. , Lim, S. K. , de Kleijn, D. P. (2013). Mesenchymal stem cell‐derived exosomes increase ATP levels, decrease oxidative stress and activate PI3K/Akt pathway to enhance myocardial viability and prevent adverse remodeling after myocardial ischemia/reperfusion injury. Stem Cell Research, 10(3), 301–312. 10.1016/j.scr.2013.01.002 - DOI - PubMed
-
- Baker, D. J. , Childs, B. G. , Durik, M. , Wijers, M. E. , Sieben, C. J. , Zhong, J. , A. Saltness, R. , Jeganathan, K. B. , Verzosa, G. C. , Pezeshki, A. , Khazaie, K. , Miller, J. D. , van Deursen, J. M. (2016). Naturally occurring p16(INK4a)‐positive cells shorten healthy lifespan. Nature, 530, 184–189. 10.1038/nature16932 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
