

Mechanism of Pacemaker Activity in Zebrafish DC2/4 Dopaminergic Neurons
- PMID: 33731451
- PMCID: PMC8176752
- DOI: 10.1523/JNEUROSCI.2124-20.2021
Mechanism of Pacemaker Activity in Zebrafish DC2/4 Dopaminergic Neurons
Abstract
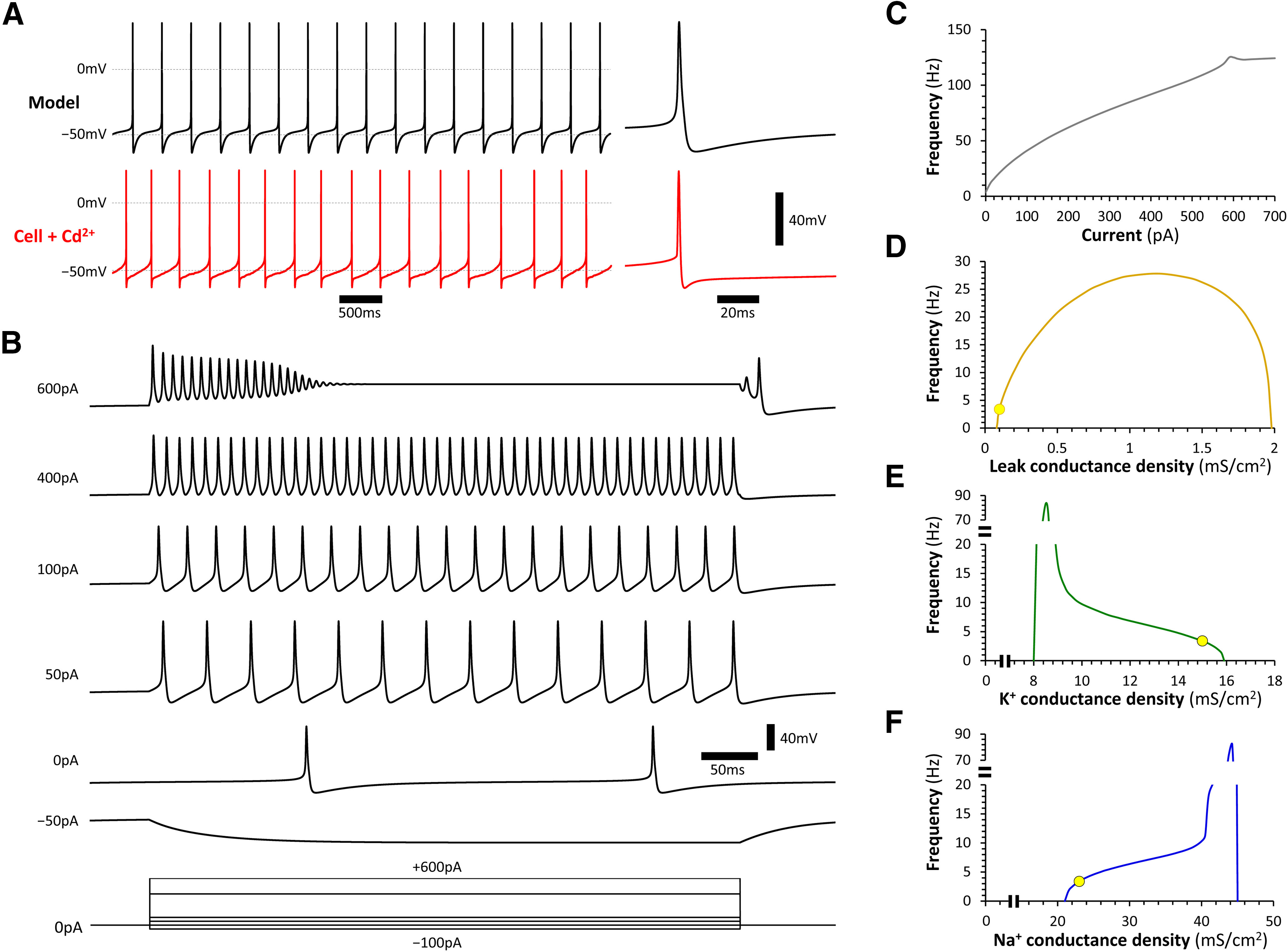
Zebrafish models are used increasingly to study the molecular pathogenesis of Parkinson's disease (PD), owing to the extensive array of techniques available for their experimental manipulation and analysis. The ascending dopaminergic projection from the posterior tuberculum (TPp; diencephalic populations DC2 and DC4) to the subpallium is considered the zebrafish correlate of the mammalian nigrostriatal projection, but little is known about the neurophysiology of zebrafish DC2/4 neurons. This is an important knowledge gap, because autonomous activity in mammalian substantia nigra (SNc) dopaminergic neurons contributes to their vulnerability in PD models. Using a new transgenic zebrafish line to label living dopaminergic neurons, and a novel brain slice preparation, we conducted whole-cell patch clamp recordings of DC2/4 neurons from adult zebrafish of both sexes. Zebrafish DC2/4 neurons share many physiological properties with mammalian dopaminergic neurons, including the cell-autonomous generation of action potentials. However, in contrast to mammalian dopaminergic neurons, the pacemaker driving intrinsic rhythmic activity in zebrafish DC2/4 neurons does not involve calcium conductances, hyperpolarization-activated cyclic nucleotide-gated (HCN) channels, or sodium leak currents. Instead, voltage clamp recordings and computational models show that interactions between three components - a small, predominantly potassium, leak conductance, voltage-gated sodium channels, and voltage-gated potassium channels - are sufficient for pacemaker activity in zebrafish DC2/4 neurons. These results contribute to understanding the comparative physiology of the dopaminergic system and provide a conceptual basis for interpreting data derived from zebrafish PD models. The findings further suggest new experimental opportunities to address the role of dopaminergic pacemaker activity in the pathogenesis of PD.SIGNIFICANCE STATEMENT Posterior tuberculum (TPp) DC2/4 dopaminergic neurons are considered the zebrafish correlate of mammalian substantia nigra (SNc) neurons, whose degeneration causes the motor signs of Parkinson's disease (PD). Our study shows that DC2/4 and SNc neurons share a number of electrophysiological properties, including depolarized membrane potential, high input resistance, and continual, cell-autonomous pacemaker activity, that strengthen the basis for the increasing use of zebrafish models to study the molecular pathogenesis of PD. The mechanisms driving pacemaker activity differ between DC2/4 and SNc neurons, providing: (1) experimental opportunities to dissociate the contributions of intrinsic activity and underlying pacemaker currents to pathogenesis; and (2) essential information for the design and interpretation of studies using zebrafish PD models.
Keywords: Parkinson's disease; computational; dopamine; electrophysiology; pacemaker; zebrafish.
Copyright © 2021 the authors.
Figures








References
-
- Banerjee K, Sinha M, Pham CLL, Jana S, Chanda D, Cappai R, Chakrabarti S (2010) Alpha-synuclein induced membrane depolarization and loss of phosphorylation capacity of isolated rat brain mitochondria: implications in Parkinson's disease. FEBS Lett 584:1571–1576. 10.1016/j.febslet.2010.03.012 - DOI - PubMed
-
- Bartel WP, Van Laar VS, Burton EA (2020) Parkinson's disease. In: Behavioral and neural genetics of zebrafish (Gerlai RT, ed), pp 377–412. San Diego: Elsevier.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous