

Human-chimpanzee fused cells reveal cis-regulatory divergence underlying skeletal evolution
- PMID: 33731941
- PMCID: PMC8038968
- DOI: 10.1038/s41588-021-00804-3
Human-chimpanzee fused cells reveal cis-regulatory divergence underlying skeletal evolution
Erratum in
-
Publisher Correction: Human-chimpanzee fused cells reveal cis-regulatory divergence underlying skeletal evolution.Nat Genet. 2021 Apr;53(4):587. doi: 10.1038/s41588-021-00849-4. Nat Genet. 2021. PMID: 33762754 No abstract available.
Abstract
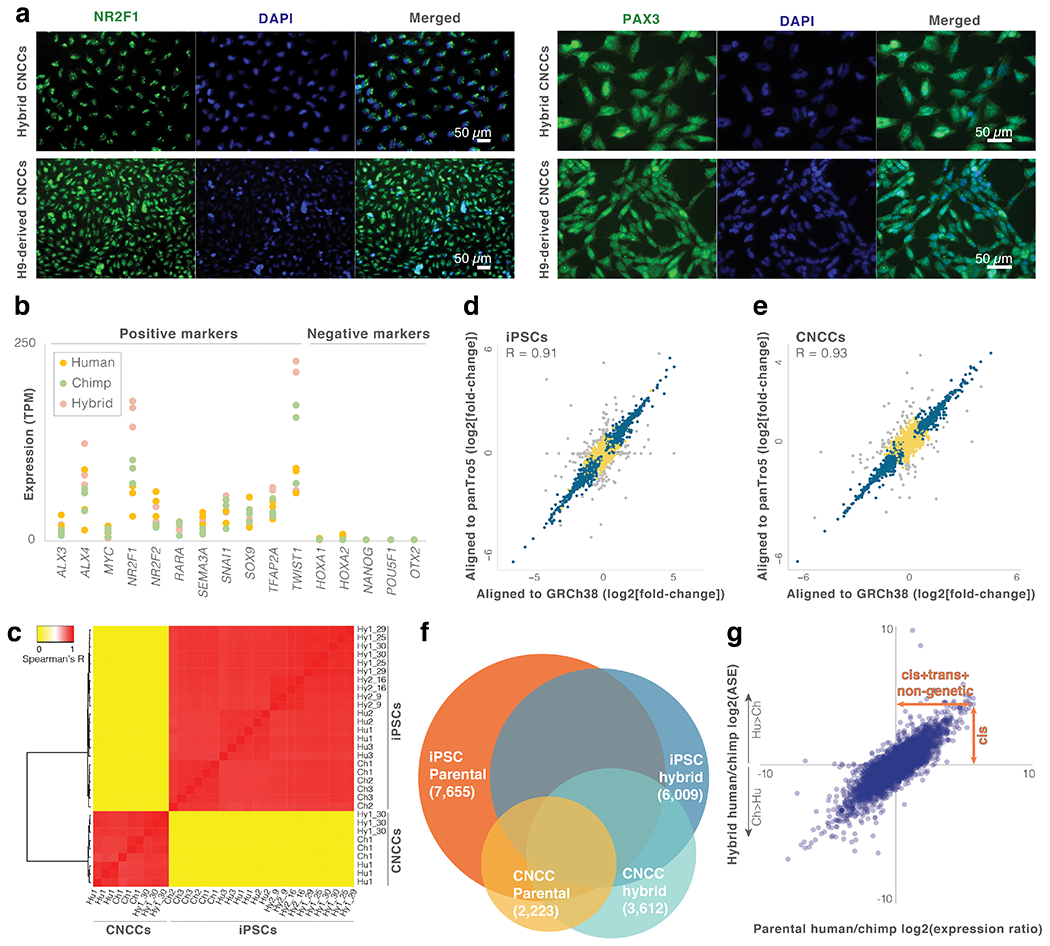
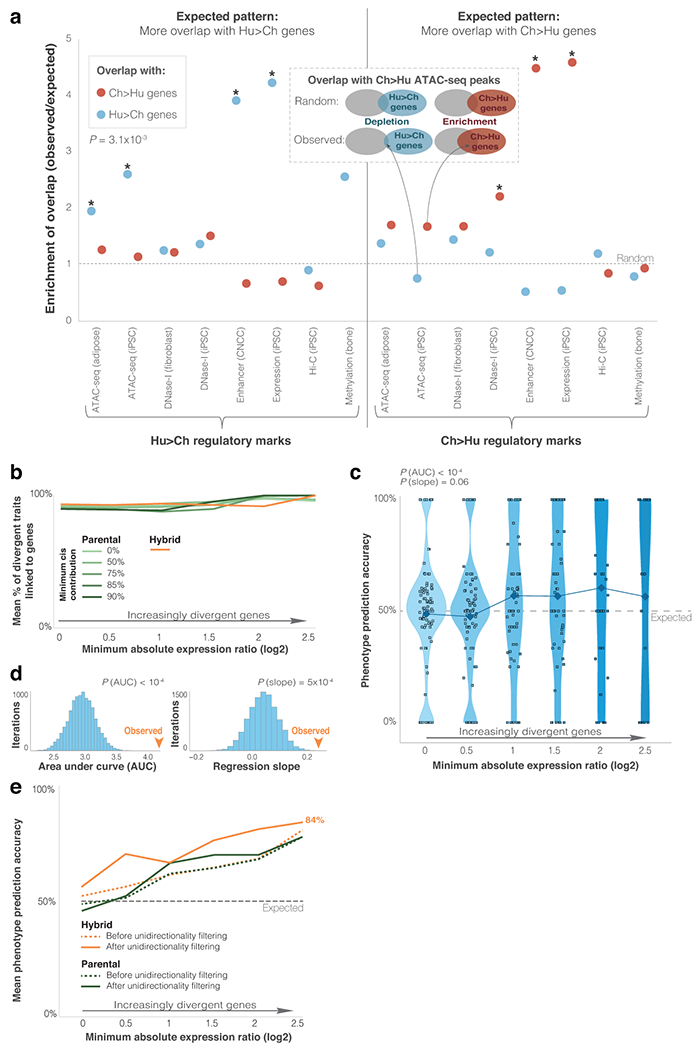
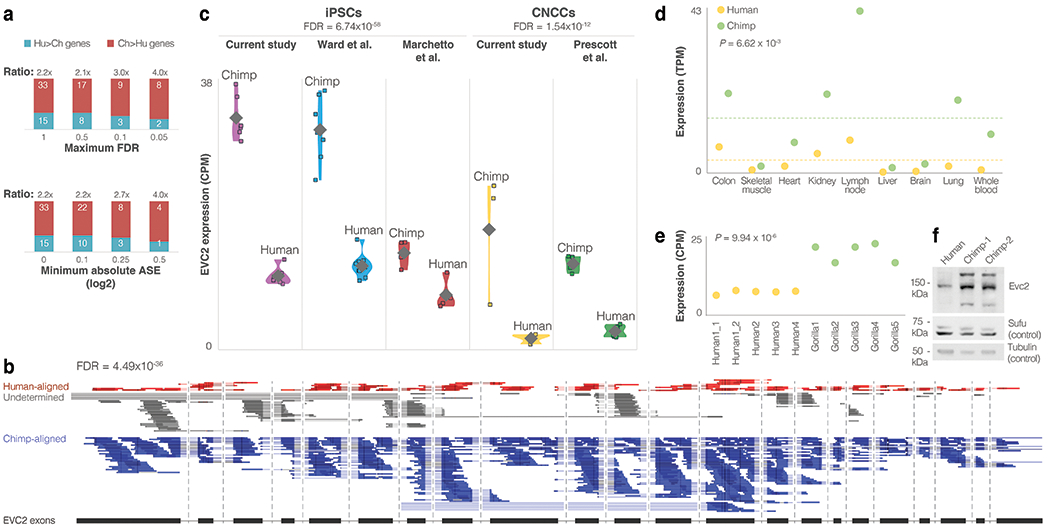
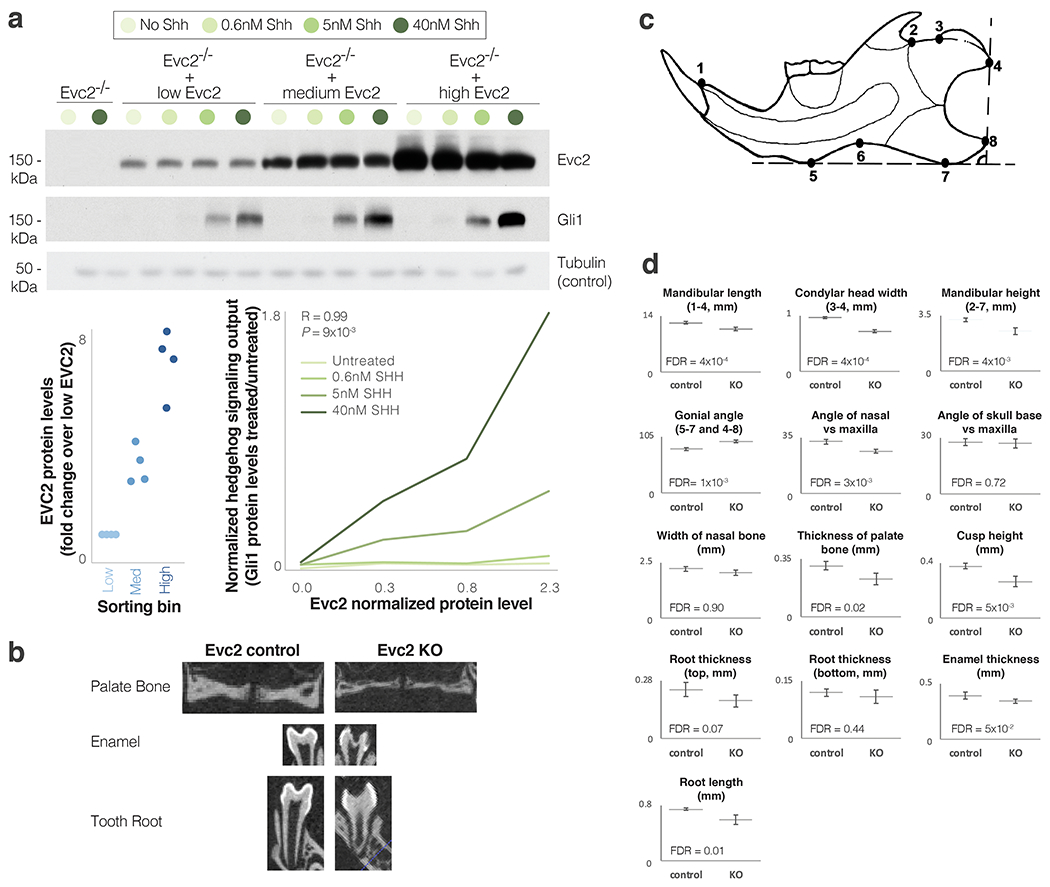
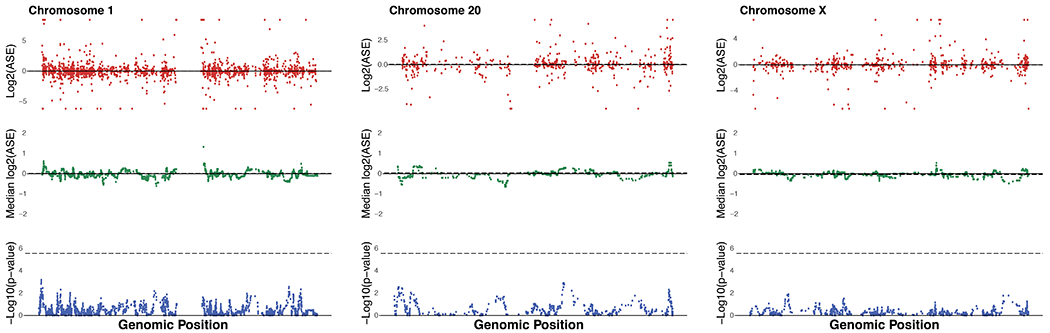
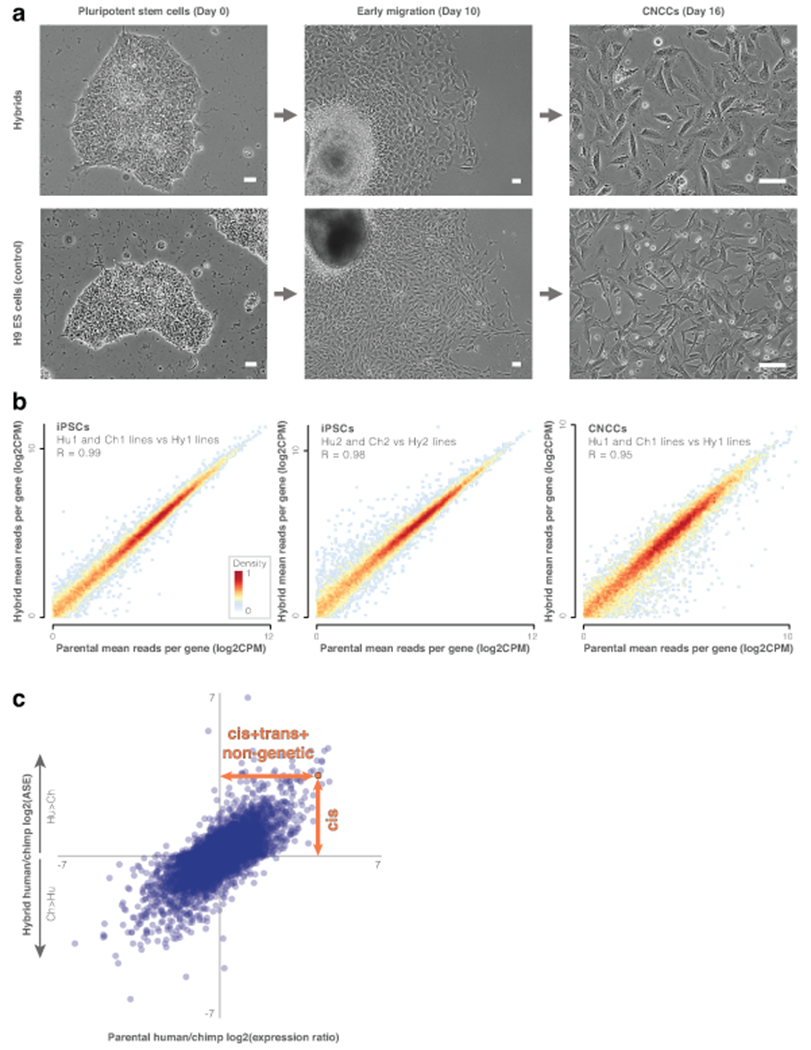
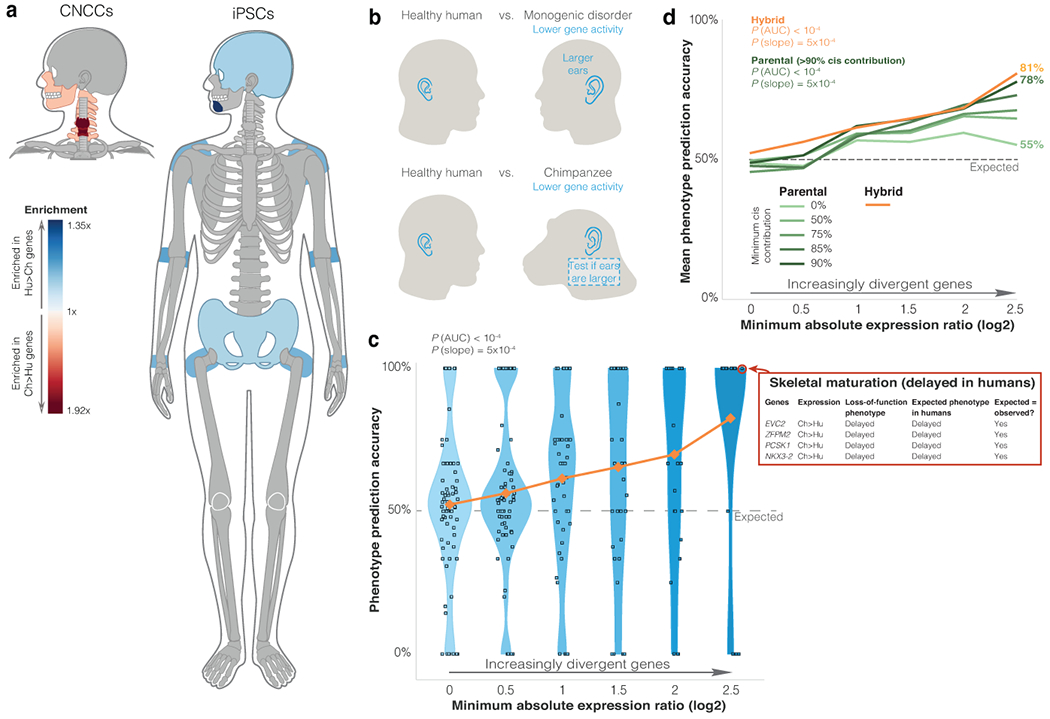
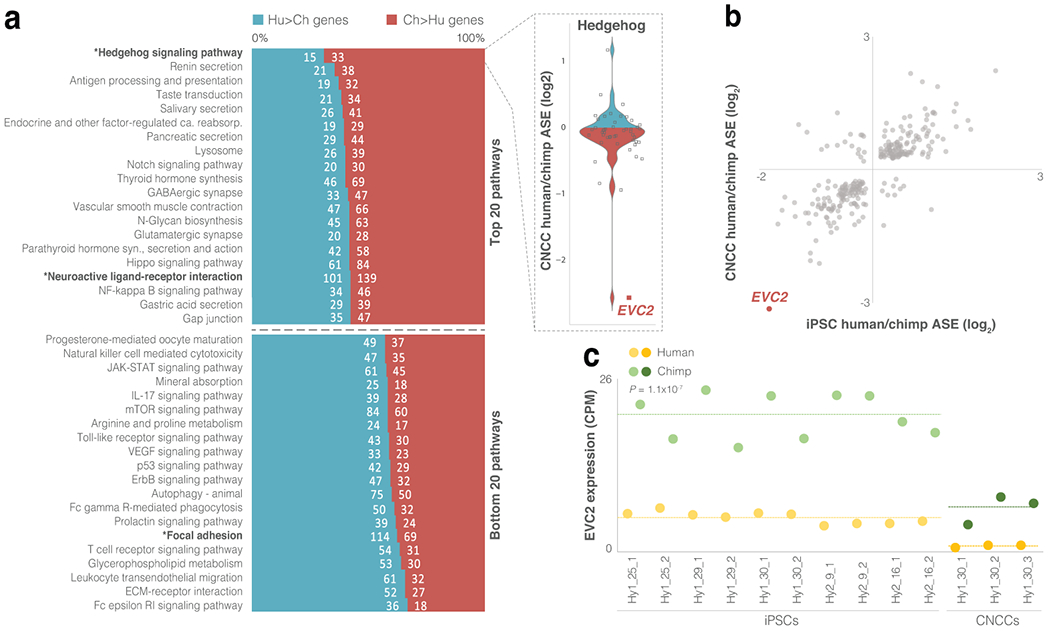
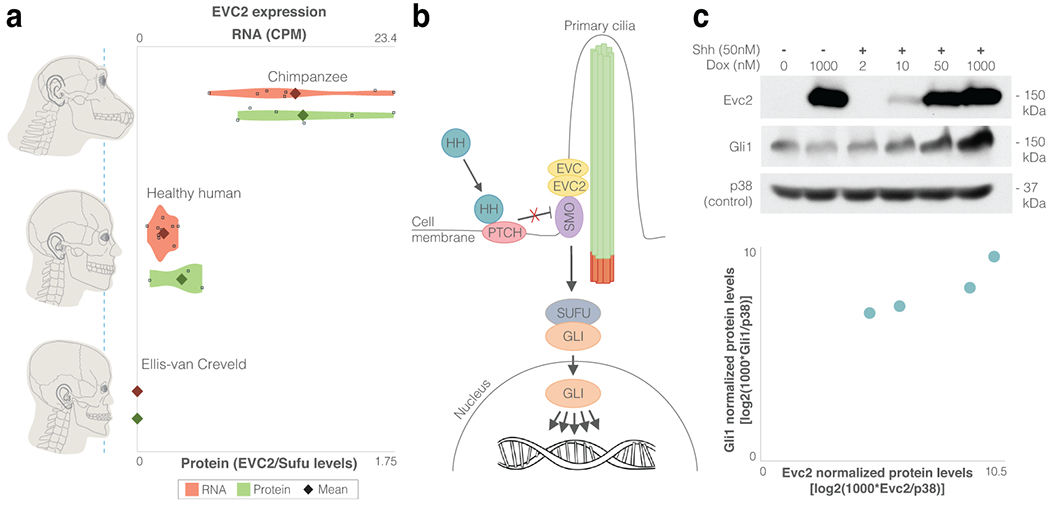
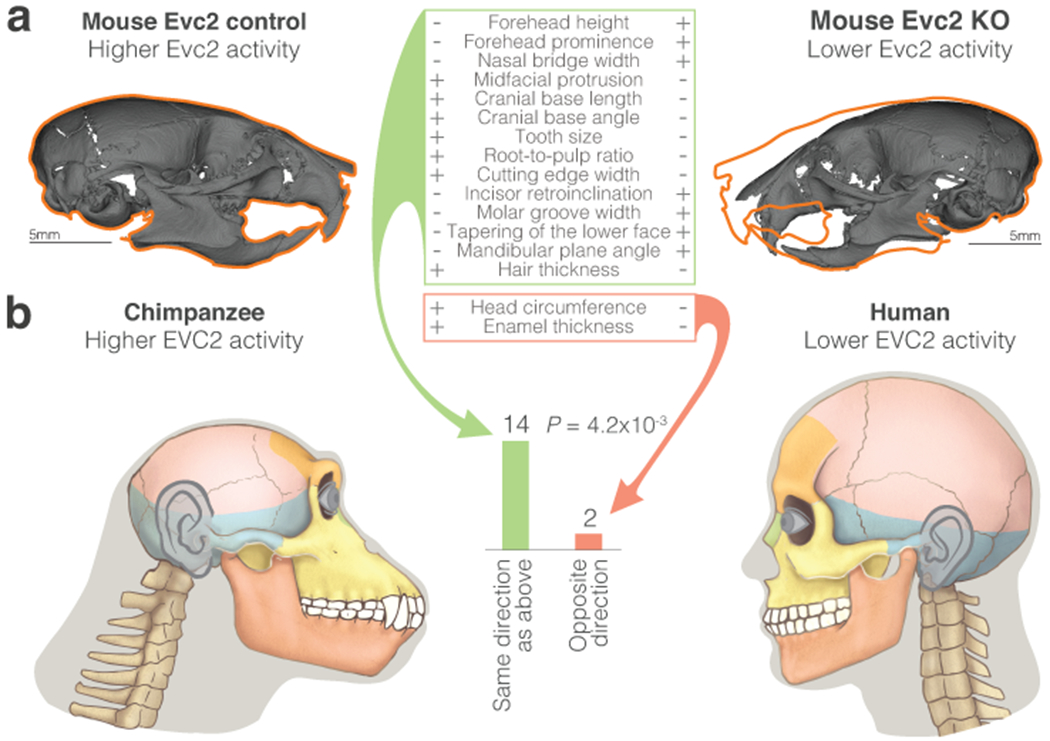
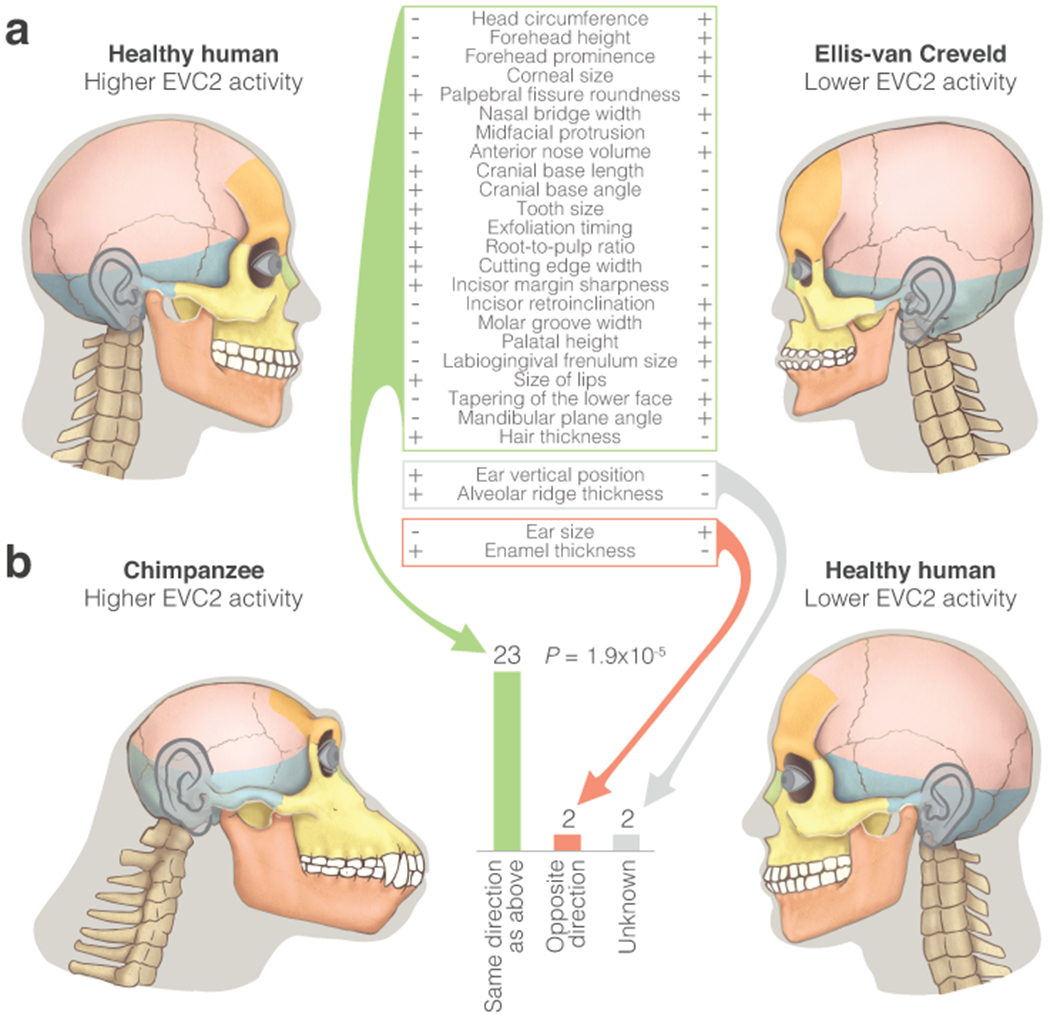
Gene regulatory divergence is thought to play a central role in determining human-specific traits. However, our ability to link divergent regulation to divergent phenotypes is limited. Here, we utilized human-chimpanzee hybrid induced pluripotent stem cells to study gene expression separating these species. The tetraploid hybrid cells allowed us to separate cis- from trans-regulatory effects, and to control for nongenetic confounding factors. We differentiated these cells into cranial neural crest cells, the primary cell type giving rise to the face. We discovered evidence of lineage-specific selection on the hedgehog signaling pathway, including a human-specific sixfold down-regulation of EVC2 (LIMBIN), a key hedgehog gene. Inducing a similar down-regulation of EVC2 substantially reduced hedgehog signaling output. Mice and humans lacking functional EVC2 show striking phenotypic parallels to human-chimpanzee craniofacial differences, suggesting that the regulatory divergence of hedgehog signaling may have contributed to the unique craniofacial morphology of humans.
Conflict of interest statement
Competing Interests Statement
The authors declare no competing interests.
Figures

Comment in
-
Studying divergence in human-chimp hybrid cells.Nat Methods. 2021 May;18(5):444. doi: 10.1038/s41592-021-01157-9. Nat Methods. 2021. PMID: 33963352 No abstract available.
References
References for main text
-
- Aiello L & Dean C An Introduction to Human Evolutionary Anatomy. (Elsevier, 2002).
-
- King MC & Wilson AC Evolution at two levels in humans and chimpanzees. Science 188, 107–116 (1975). - PubMed
Methods-only References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
