

Local Homeostatic Regulation of the Spectral Radius of Echo-State Networks
- PMID: 33732127
- PMCID: PMC7958921
- DOI: 10.3389/fncom.2021.587721
Local Homeostatic Regulation of the Spectral Radius of Echo-State Networks
Abstract
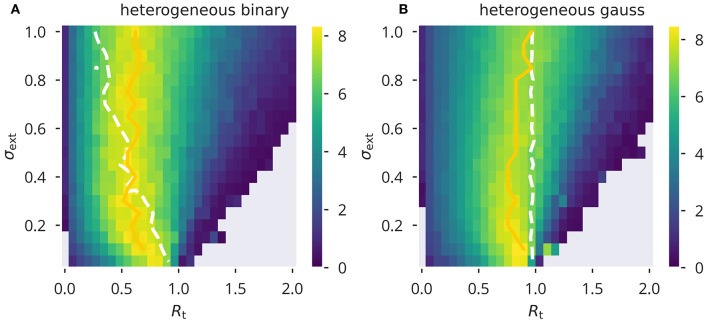
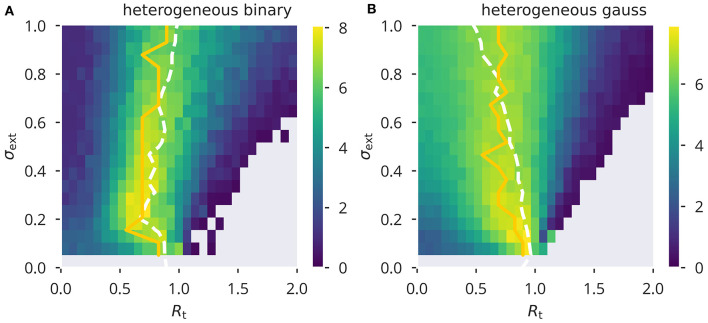
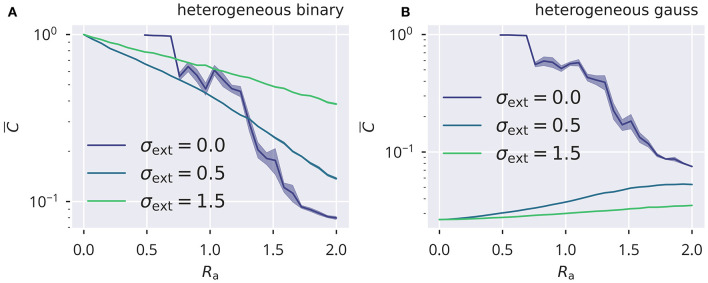
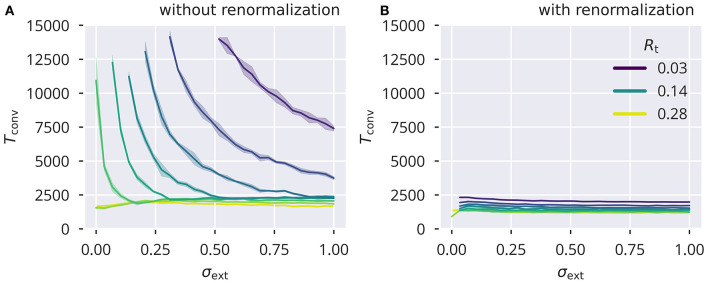
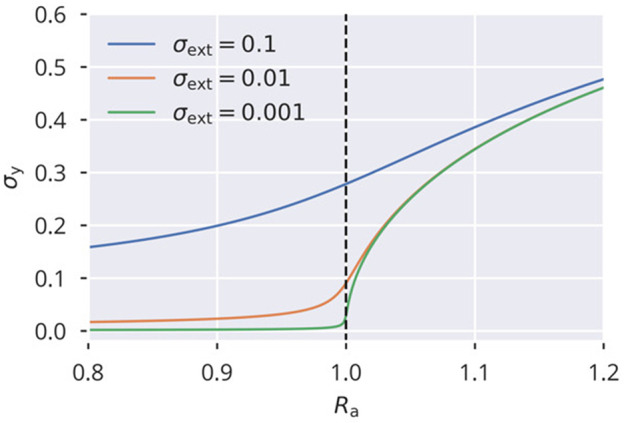
Recurrent cortical networks provide reservoirs of states that are thought to play a crucial role for sequential information processing in the brain. However, classical reservoir computing requires manual adjustments of global network parameters, particularly of the spectral radius of the recurrent synaptic weight matrix. It is hence not clear if the spectral radius is accessible to biological neural networks. Using random matrix theory, we show that the spectral radius is related to local properties of the neuronal dynamics whenever the overall dynamical state is only weakly correlated. This result allows us to introduce two local homeostatic synaptic scaling mechanisms, termed flow control and variance control, that implicitly drive the spectral radius toward the desired value. For both mechanisms the spectral radius is autonomously adapted while the network receives and processes inputs under working conditions. We demonstrate the effectiveness of the two adaptation mechanisms under different external input protocols. Moreover, we evaluated the network performance after adaptation by training the network to perform a time-delayed XOR operation on binary sequences. As our main result, we found that flow control reliably regulates the spectral radius for different types of input statistics. Precise tuning is however negatively affected when interneural correlations are substantial. Furthermore, we found a consistent task performance over a wide range of input strengths/variances. Variance control did however not yield the desired spectral radii with the same precision, being less consistent across different input strengths. Given the effectiveness and remarkably simple mathematical form of flow control, we conclude that self-consistent local control of the spectral radius via an implicit adaptation scheme is an interesting and biological plausible alternative to conventional methods using set point homeostatic feedback controls of neural firing.
Keywords: echo-state networks; homeostasis; recurrent networks; reservoir computing; spectral radius; synaptic scaling.
Copyright © 2021 Schubert and Gros.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures





References
-
- Abbott L. F., LeMasson G. (1993). Analysis of neuron models with dynamically regulated conductances. Neural Comput. 5, 823–842. 10.1162/neco.1993.5.6.823 - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials