

Impact of serine protease inhibitor alpha1-antitrypsin on expression of endoplasmic reticulum stress-induced proinflammatory factors in adipocytes
- PMID: 33732904
- PMCID: PMC7937654
- DOI: 10.1016/j.bbrep.2021.100967
Impact of serine protease inhibitor alpha1-antitrypsin on expression of endoplasmic reticulum stress-induced proinflammatory factors in adipocytes
Abstract
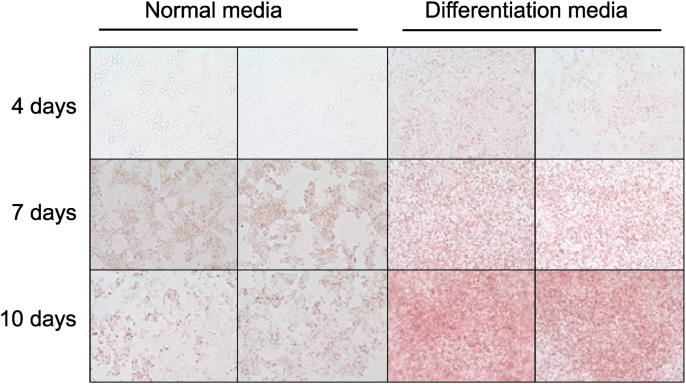
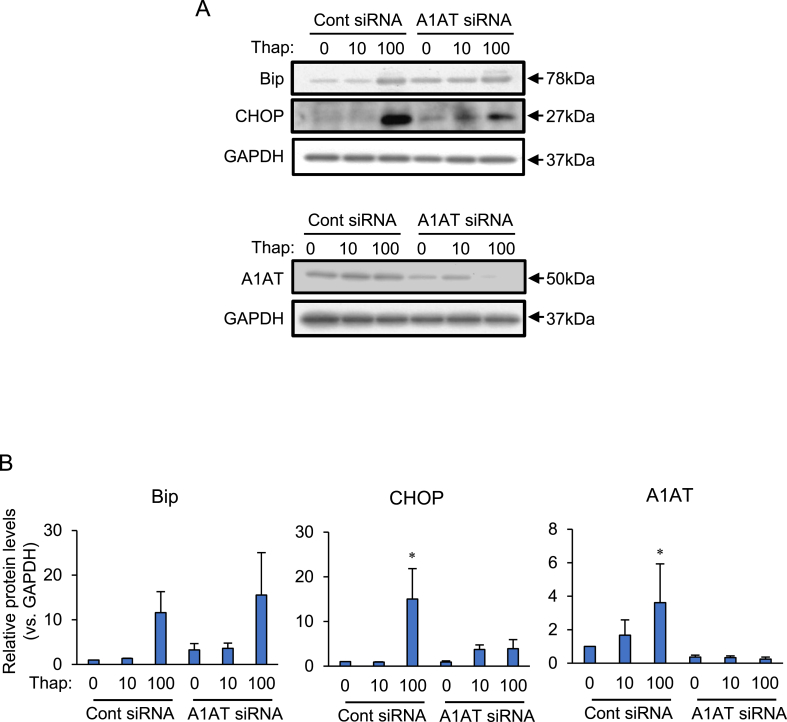
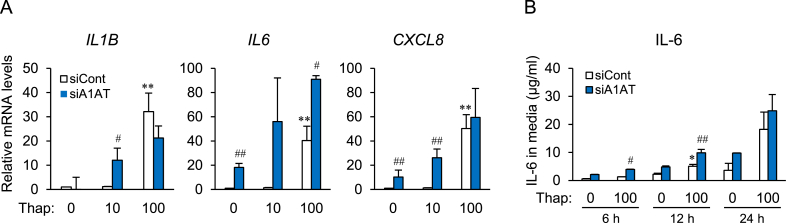
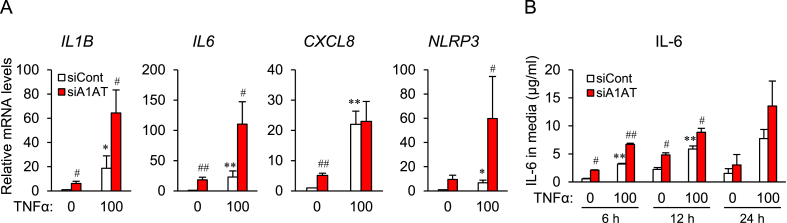
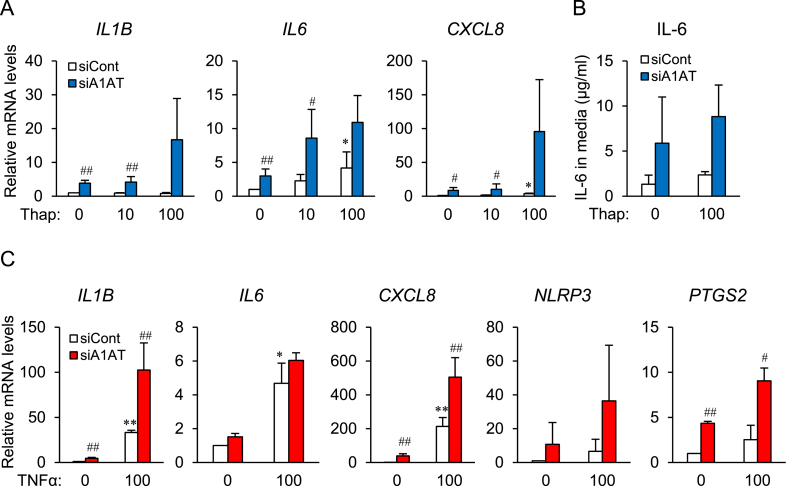
Obesity-induced endoplasmic reticulum (ER) stress contributes to low-grade chronic inflammation in adipose tissue and may cause metabolic disorders such as diabetes mellitus and dyslipidemia. Identification of high serpina A1 (alpha-1 antitrypsin, A1AT) expression in mouse adipose tissue and adipocytes prompted us to explore the role of A1AT in the inflammatory response of adipocytes under ER stress. We aimed to determine the role of A1AT expression in adipocytes with ER stress during regulation of adipocyte homeostasis and inflammation. To this end, we chemically induced ER stress in A1AT small interfering RNA-transfected differentiating adipocytes using thapsigargin. Induction of CCAAT-enhancer-binding protein homologous protein (CHOP), an ER stress marker, by thapsigargin was lower in A1AT-deficient SW872 adipocytes. Thapsigargin or the proinflammatory cytokine tumor necrosis factor (TNF)α increased basal expression of cytokines such as interleukin (IL)-1β and IL-8 in both SW872 and primary omental adipocytes. This thapsigargin- or TNFα-induced expression of proinflammatory genes was increased by A1AT deficiency. These findings indicate that adipose A1AT may suppress the ER stress response to block excessive expression of proinflammatory factors, which suggests that A1AT protects against adipose tissue dysfunction associated with ER stress activation.
Keywords: Adipocyte; Alpha-1 antitrypsin; Endoplasmic reticulum stress; Proinflammatory factor.
© 2021 Published by Elsevier B.V.
Conflict of interest statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
Similar articles
-
Alpha-1 Antitrypsin-Induced Endoplasmic Reticulum Stress Promotes Invasion by Extravillous Trophoblasts.Int J Mol Sci. 2021 Apr 1;22(7):3683. doi: 10.3390/ijms22073683. Int J Mol Sci. 2021. PMID: 33916165 Free PMC article.
-
Effect of endoplasmic reticulum stress on inflammation and adiponectin regulation in human adipocytes.Metab Syndr Relat Disord. 2012 Aug;10(4):297-306. doi: 10.1089/met.2012.0002. Epub 2012 Apr 30. Metab Syndr Relat Disord. 2012. PMID: 22545589 Free PMC article.
-
Aquaporin-11 Contributes to TGF-β1-Induced Endoplasmic Reticulum Stress in Human Visceral Adipocytes: Role in Obesity-Associated Inflammation.Cells. 2020 Jun 4;9(6):1403. doi: 10.3390/cells9061403. Cells. 2020. PMID: 32512939 Free PMC article.
-
The Pathogenesis of Obesity-Associated Adipose Tissue Inflammation.Adv Exp Med Biol. 2017;960:221-245. doi: 10.1007/978-3-319-48382-5_9. Adv Exp Med Biol. 2017. PMID: 28585201 Review.
-
Alpha-1-antitrypsin deficiency: current concepts.Lung. 2007 Jul-Aug;185(4):191-201. doi: 10.1007/s00408-007-9009-y. Epub 2007 Jun 12. Lung. 2007. PMID: 17562108 Review.
Cited by
-
Influence of Protein Carbonylation on Human Adipose Tissue Dysfunction in Obesity and Insulin Resistance.Biomedicines. 2022 Nov 24;10(12):3032. doi: 10.3390/biomedicines10123032. Biomedicines. 2022. PMID: 36551793 Free PMC article.
-
Intermittent Cold Exposure Induces Distinct Proteomic Signatures in White Adipose Tissue of Mice.Int J Mol Sci. 2025 Aug 15;26(16):7898. doi: 10.3390/ijms26167898. Int J Mol Sci. 2025. PMID: 40869217 Free PMC article.
-
Alpha-1 Antitrypsin-Induced Endoplasmic Reticulum Stress Promotes Invasion by Extravillous Trophoblasts.Int J Mol Sci. 2021 Apr 1;22(7):3683. doi: 10.3390/ijms22073683. Int J Mol Sci. 2021. PMID: 33916165 Free PMC article.
-
Effect of Cordyceps militaris extract containing cordycepin on the adipogenesis and lipolysis of adipocytes.FEBS Open Bio. 2025 Feb;15(2):335-345. doi: 10.1002/2211-5463.13930. Epub 2024 Nov 21. FEBS Open Bio. 2025. PMID: 39572891 Free PMC article.
-
Role of Transcription Factor, LIM Homeobox 9 (LHX9) in Inflammatory Response by PGE2 and Thrombin in SERPINA1-Silencing Endometrial Stromal Cells.Mol Reprod Dev. 2025 Aug;92(8):e70046. doi: 10.1002/mrd.70046. Mol Reprod Dev. 2025. PMID: 40802848 Free PMC article.
References
-
- Martin G.S., Mannino D.M., Eaton S., Moss M. The epidemiology of sepsis in the United States from 1979 through 2000. N. Engl. J. Med. 2003;348:1546–1554. - PubMed
-
- Matsuda A., Jacob A., Wu R., Aziz M., Yang W.L., Matsutani T., Suzuki H., Furukawa K., Uchida E., Wang Novel therapeutic targets for sepsis: regulation of exaggerated inflammatory responses. J. Nippon Med. Sch. 2012;79:4–18. - PubMed
-
- Singer M., Deutschman C.S., Seymour C.W., Shankar-Hari M., Annane D., Bauer M., Bellomo R., Bernard G.R., Chiche J.D., Coopersmith C.M., Hotchkiss R.S., Levy M.M., Marshall J.C., Martin G.S., Opal S.M., Rubenfeld G.D., van der Poll T., Vincent J.L., Angus The third international consensus definitions for sepsis and septic shock (Sepsis-3) J. Am. Med. Assoc. 2016;315:801–810. - PMC - PubMed
-
- Redl H., Schlag G., Bahrami S. Animal models of sepsis and shock: a review and lessons learned. Edwin A Deith. Shock. 1998;10:442–445. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
