

Energy metabolism design of the striated muscle cell
- PMID: 33733879
- PMCID: PMC8576364
- DOI: 10.1152/physrev.00040.2020
Energy metabolism design of the striated muscle cell
Abstract
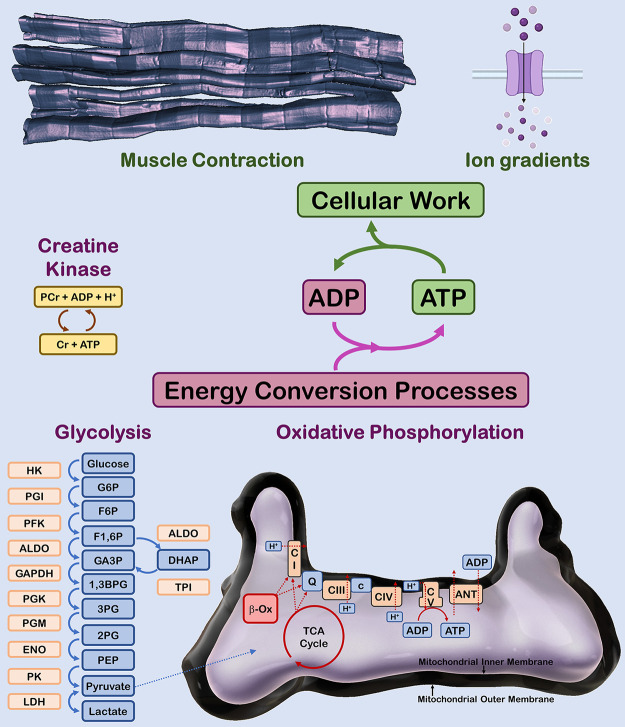
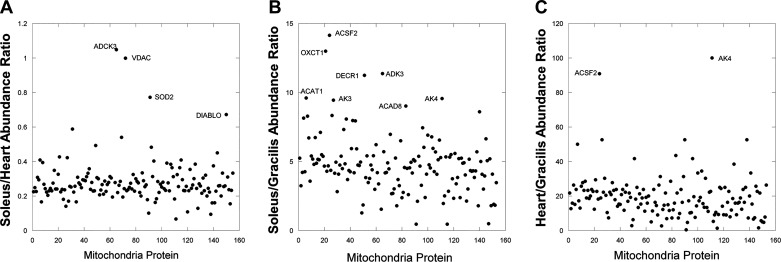
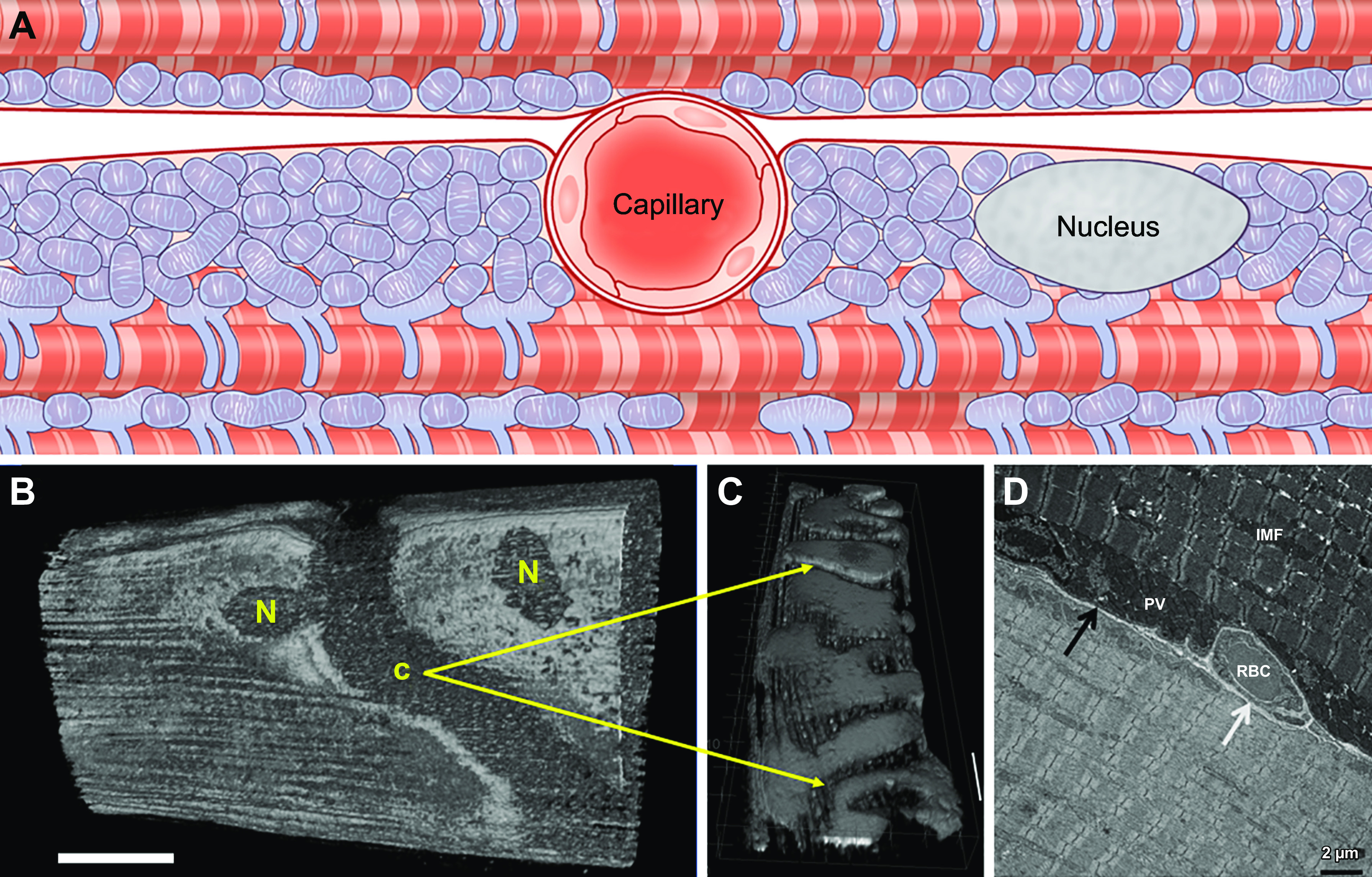
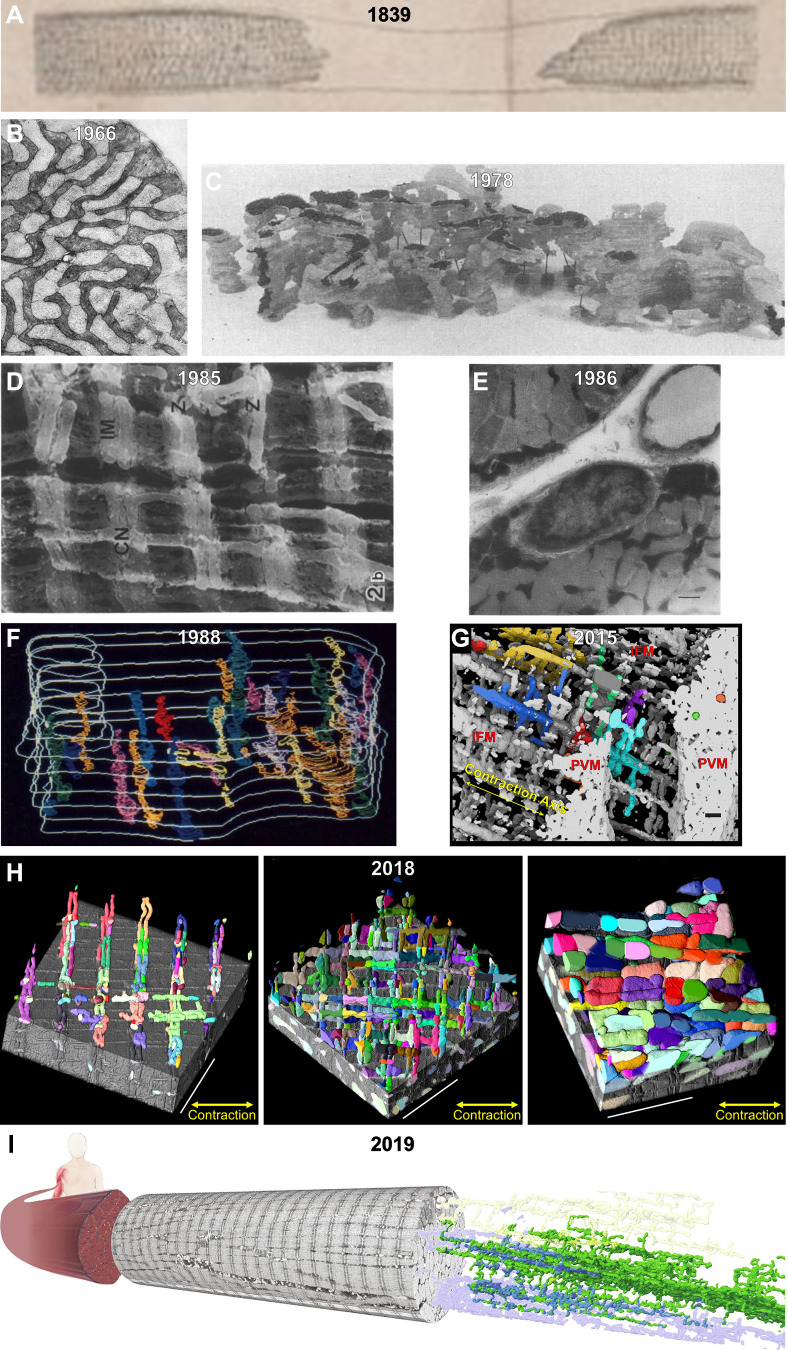
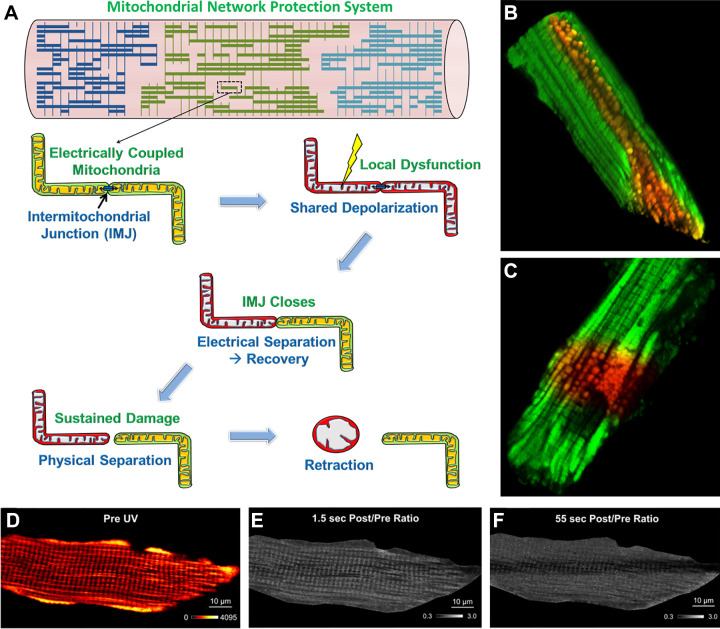
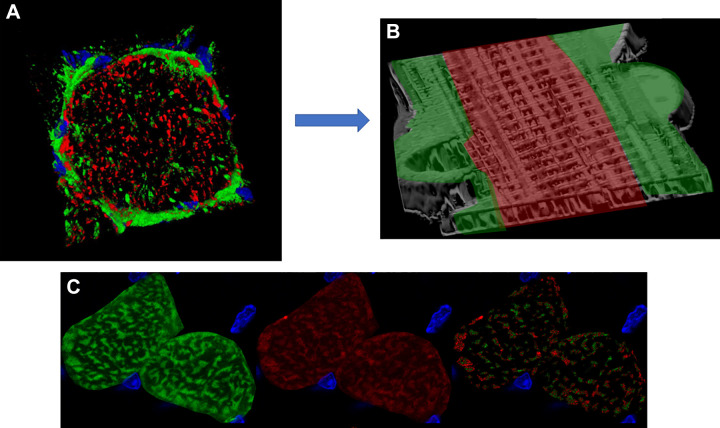
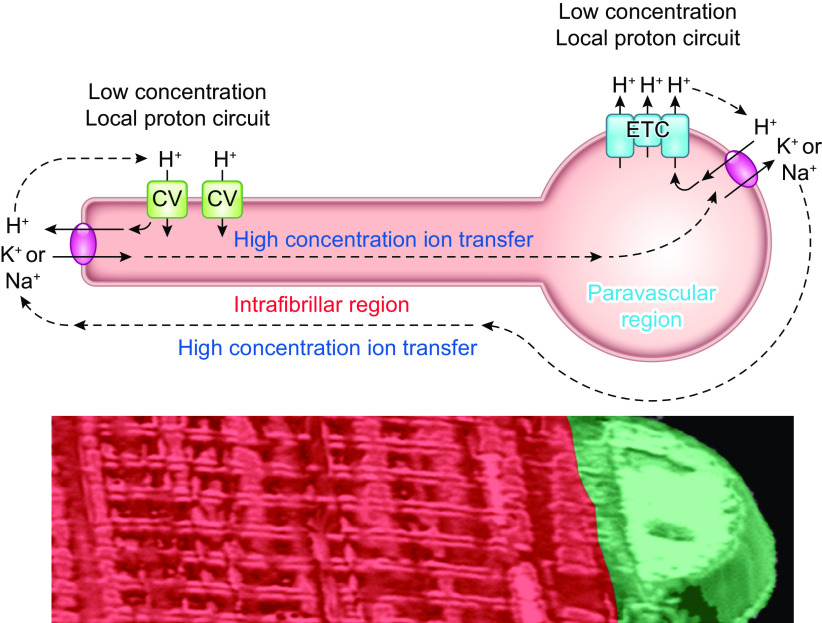
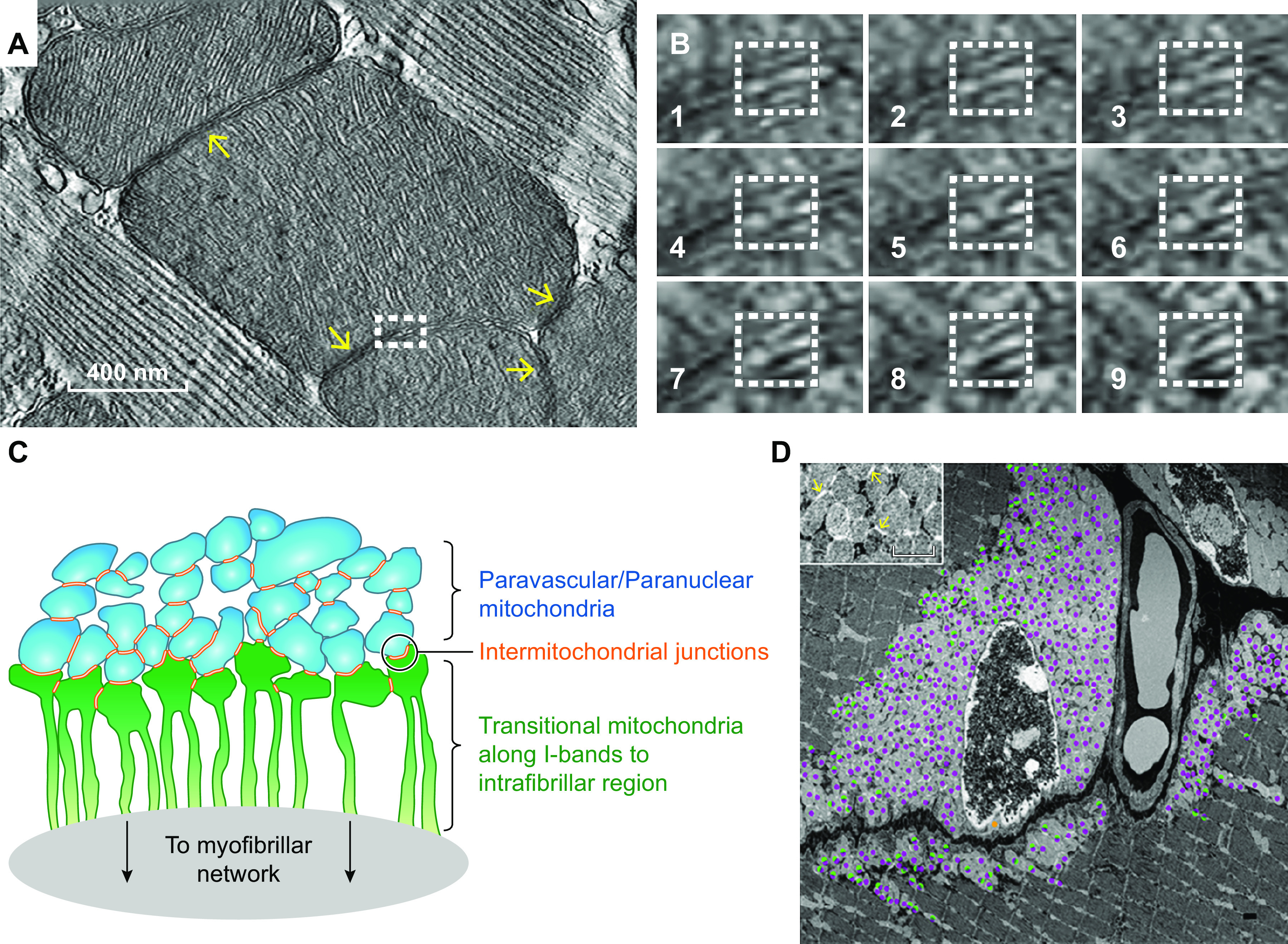
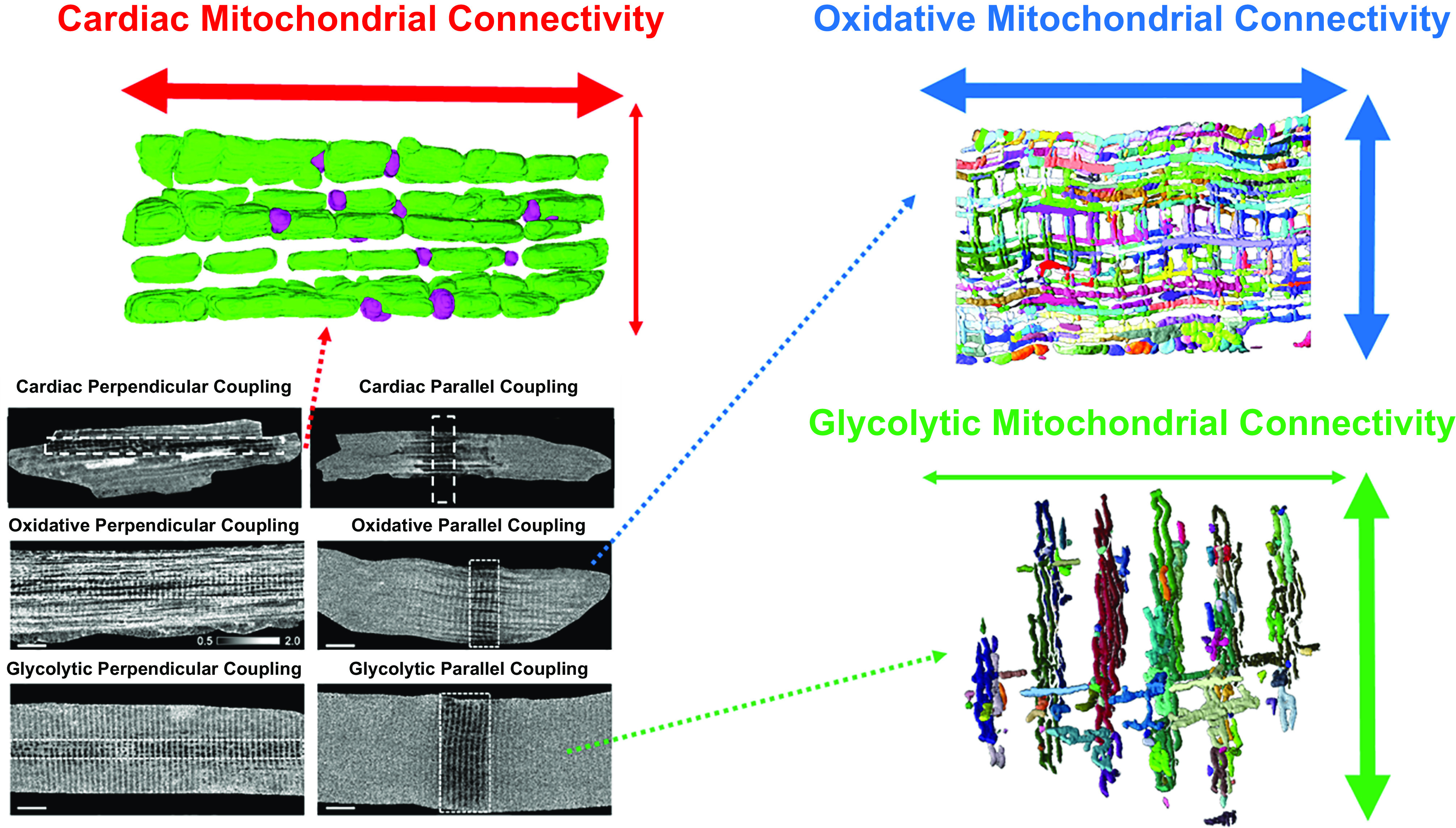
The design of the energy metabolism system in striated muscle remains a major area of investigation. Here, we review our current understanding and emerging hypotheses regarding the metabolic support of muscle contraction. Maintenance of ATP free energy, so called energy homeostasis, via mitochondrial oxidative phosphorylation is critical to sustained contractile activity, and this major design criterion is the focus of this review. Cell volume invested in mitochondria reduces the space available for generating contractile force, and this spatial balance between mitochondria acontractile elements to meet the varying sustained power demands across muscle types is another important design criterion. This is accomplished with remarkably similar mass-specific mitochondrial protein composition across muscle types, implying that it is the organization of mitochondria within the muscle cell that is critical to supporting sustained muscle function. Beyond the production of ATP, ubiquitous distribution of ATPases throughout the muscle requires rapid distribution of potential energy across these large cells. Distribution of potential energy has long been thought to occur primarily through facilitated metabolite diffusion, but recent analysis has questioned the importance of this process under normal physiological conditions. Recent structural and functional studies have supported the hypothesis that the mitochondrial reticulum provides a rapid energy distribution system via the conduction of the mitochondrial membrane potential to maintain metabolic homeostasis during contractile activity. We extensively review this aspect of the energy metabolism design contrasting it with metabolite diffusion models and how mitochondrial structure can play a role in the delivery of energy in the striated muscle.
Keywords: cellular energy distribution; mitochondria; mitochondrial networks; mitochondrial reticulum; oxidative phosphorylation.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the authors.
Figures


References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources