

Direct unfolding of RuvA-HJ complex at the single-molecule level
- PMID: 33737156
- PMCID: PMC8204333
- DOI: 10.1016/j.bpj.2021.03.006
Direct unfolding of RuvA-HJ complex at the single-molecule level
Abstract
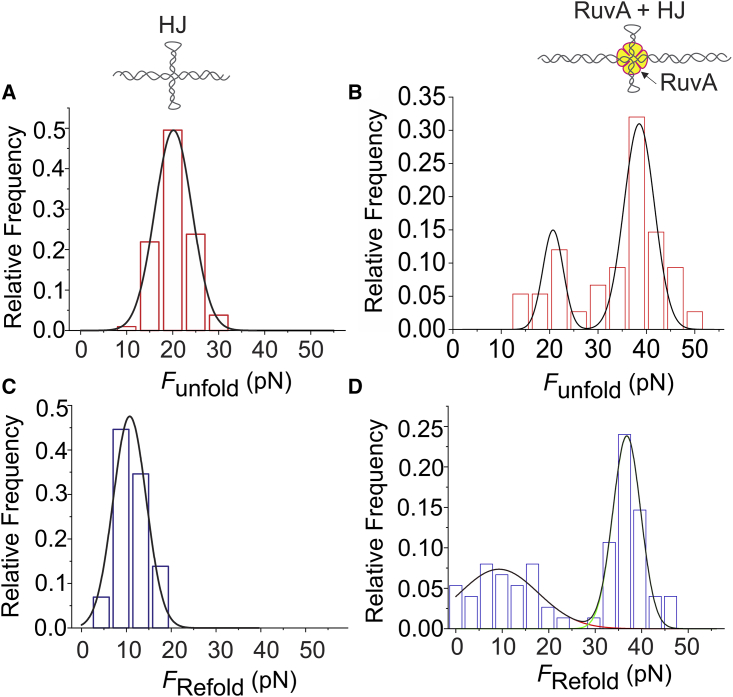
The repair of double-stranded DNA breaks via homologous recombination involves a four-way cross-strand intermediate known as Holliday junction (HJ), which is recognized, processed, and resolved by a specific set of proteins. RuvA, a prokaryotic HJ-binding protein, is known to stabilize the square-planar conformation of the HJ, which is otherwise a short-lived intermediate. Despite much progress being made regarding the molecular mechanism of RuvA-HJ interactions, the mechanochemical aspect of this protein-HJ complex is yet to be investigated. Here, we employed an optical-tweezers-based, single-molecule manipulation assay to detect the formation of RuvA-HJ complex and determined its mechanical and thermodynamic properties in a manner that would be impossible with traditional ensemble techniques. We found that the binding of RuvA increases the unfolding force (Funfold) of the HJ by ∼2-fold. Compared with the ΔGunfold of the HJ alone (54 ± 13 kcal/mol), the increased free energy of the RuvA-HJ complex (101 ± 20 kcal/mol) demonstrates that the RuvA protein stabilizes HJs. Interestingly, the protein remains bound to the mechanically melted HJ, facilitating its refolding at an unusually high force when the stretched DNA molecule is relaxed. These results suggest that the RuvA protein not only stabilizes the HJs but also induces refolding of the HJs. The single-molecule platform that we employed here for studying the RuvA-HJ interaction is broadly applicable to study other HJ-binding proteins involved in the critical DNA repair process.
Copyright © 2021 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures




Similar articles
-
Single-Molecule Imaging Reveals Conformational Manipulation of Holliday Junction DNA by the Junction Processing Protein RuvA.Biochemistry. 2018 Jul 3;57(26):3616-3624. doi: 10.1021/acs.biochem.8b00404. Epub 2018 May 24. Biochemistry. 2018. PMID: 29767969
-
The RuvA homologues from Mycoplasma genitalium and Mycoplasma pneumoniae exhibit unique functional characteristics.PLoS One. 2012;7(5):e38301. doi: 10.1371/journal.pone.0038301. Epub 2012 May 30. PLoS One. 2012. PMID: 22666500 Free PMC article.
-
Formation of a stable RuvA protein double tetramer is required for efficient branch migration in vitro and for replication fork reversal in vivo.J Biol Chem. 2011 Jun 24;286(25):22372-83. doi: 10.1074/jbc.M111.233908. Epub 2011 Apr 29. J Biol Chem. 2011. PMID: 21531731 Free PMC article.
-
Homologous Recombination under the Single-Molecule Fluorescence Microscope.Int J Mol Sci. 2019 Dec 3;20(23):6102. doi: 10.3390/ijms20236102. Int J Mol Sci. 2019. PMID: 31816946 Free PMC article. Review.
-
Properties of the Escherichia coli RuvA and RuvB proteins involved in DNA repair, recombination and mutagenesis.Biochimie. 1991 Apr;73(4):505-7. doi: 10.1016/0300-9084(91)90120-p. Biochimie. 1991. PMID: 1911951 Review.
Cited by
-
Search and processing of Holliday junctions within long DNA by junction-resolving enzymes.Nat Commun. 2022 Oct 7;13(1):5921. doi: 10.1038/s41467-022-33503-6. Nat Commun. 2022. PMID: 36207294 Free PMC article.
-
Measuring the biomechanical properties of cell-derived fibronectin fibrils.Biomech Model Mechanobiol. 2025 Apr;24(2):455-469. doi: 10.1007/s10237-024-01918-3. Epub 2024 Dec 26. Biomech Model Mechanobiol. 2025. PMID: 39725835 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
