

Curvature sensing amphipathic helix in the C-terminus of RTNLB13 is conserved in all endoplasmic reticulum shaping reticulons in Arabidopsis thaliana
- PMID: 33737685
- PMCID: PMC7973432
- DOI: 10.1038/s41598-021-85866-3
Curvature sensing amphipathic helix in the C-terminus of RTNLB13 is conserved in all endoplasmic reticulum shaping reticulons in Arabidopsis thaliana
Abstract
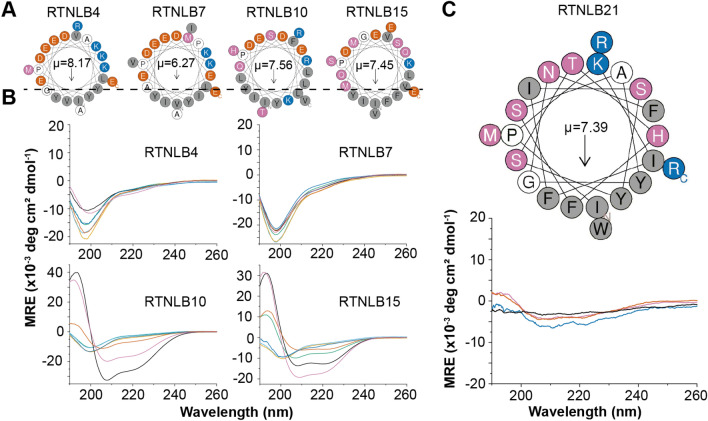
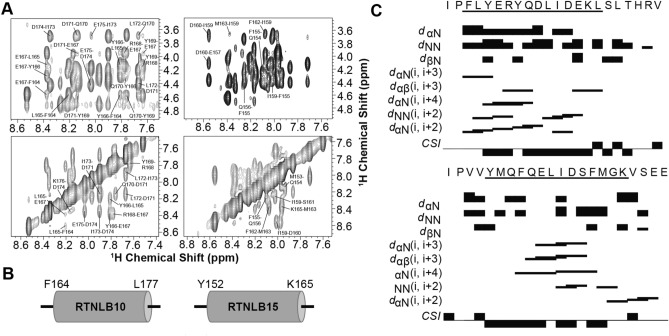
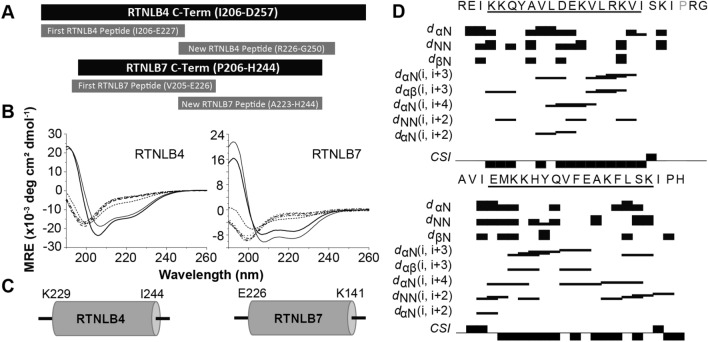
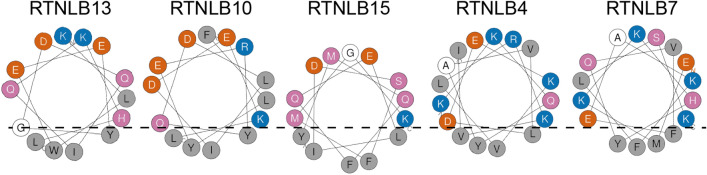
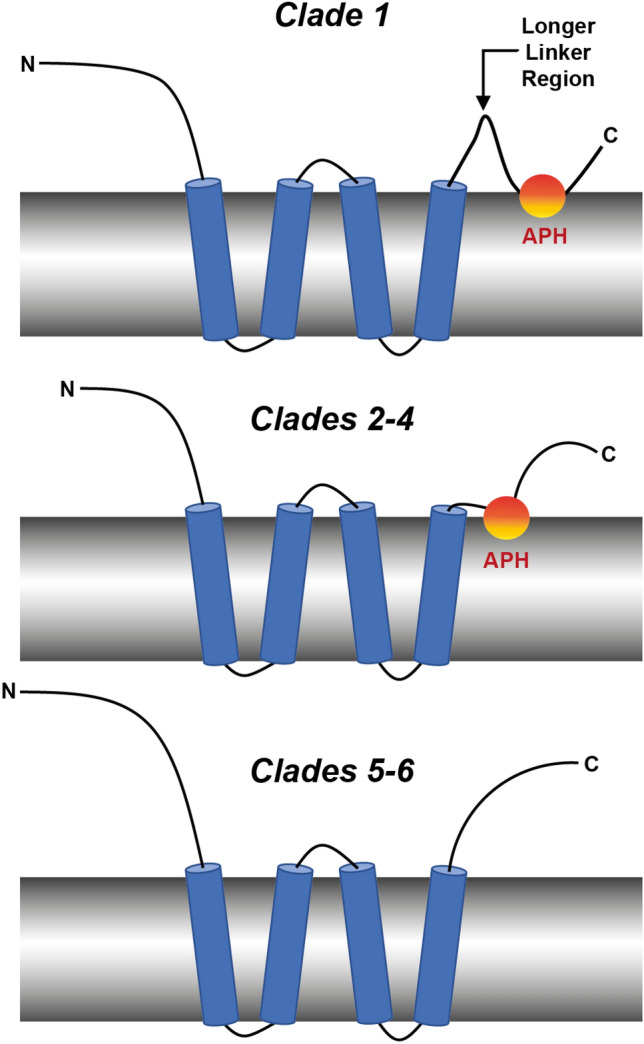
The reticulon family of integral membrane proteins are conserved across all eukaryotes and typically localize to the endoplasmic reticulum (ER), where they are involved in generating highly-curved tubules. We recently demonstrated that Reticulon-like protein B13 (RTNLB13) from Arabidopsis thaliana contains a curvature-responsive amphipathic helix (APH) important for the proteins' ability to induce curvature in the ER membrane, but incapable of generating curvature by itself. We suggested it acts as a feedback element, only folding/binding once a sufficient degree of curvature has been achieved, and stabilizes curvature without disrupting the bilayer. However, it remains unclear whether this is unique to RTNLB13 or is conserved across all reticulons-to date, experimental evidence has only been reported for two reticulons. Here we used biophysical methods to characterize a minimal library of putative APH peptides from across the 21 A. thaliana isoforms. We found that reticulons with the closest evolutionary relationship to RTNLB13 contain curvature-sensing APHs in the same location with sequence conservation. Our data reveal that a more distantly-related branch of reticulons developed a ~ 20-residue linker between the transmembrane domain and APH. This may facilitate functional flexibility as previous studies have linked these isoforms not only to ER remodeling but other cellular activities.
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Revealing the mechanism of protein-lipid interactions for a putative membrane curvature sensor in plant endoplasmic reticulum.Biochim Biophys Acta Biomembr. 2020 Mar 1;1862(3):183160. doi: 10.1016/j.bbamem.2019.183160. Epub 2019 Dec 24. Biochim Biophys Acta Biomembr. 2020. PMID: 31874147
-
Five Arabidopsis reticulon isoforms share endoplasmic reticulum location, topology, and membrane-shaping properties.Plant Cell. 2010 Apr;22(4):1333-43. doi: 10.1105/tpc.110.074385. Epub 2010 Apr 27. Plant Cell. 2010. PMID: 20424177 Free PMC article.
-
Bacterial expression, purification and biophysical characterization of the smallest plant reticulon isoform, RTNLB13.Protein Expr Purif. 2018 Dec;152:31-39. doi: 10.1016/j.pep.2018.06.015. Epub 2018 Jun 30. Protein Expr Purif. 2018. PMID: 29969670
-
Endoplasmatic reticulum shaping by generic mechanisms and protein-induced spontaneous curvature.Adv Colloid Interface Sci. 2014 Jun;208:153-60. doi: 10.1016/j.cis.2014.02.006. Epub 2014 Feb 15. Adv Colloid Interface Sci. 2014. PMID: 24607001 Review.
-
The reticulons: guardians of the structure and function of the endoplasmic reticulum.Exp Cell Res. 2012 Jul 1;318(11):1201-7. doi: 10.1016/j.yexcr.2012.03.002. Epub 2012 Mar 9. Exp Cell Res. 2012. PMID: 22425683 Review.
Cited by
-
Evolution of factors shaping the endoplasmic reticulum.Traffic. 2022 Sep;23(9):462-473. doi: 10.1111/tra.12863. Epub 2022 Aug 17. Traffic. 2022. PMID: 36040076 Free PMC article.
-
RTN family and its response to abiotic stress in Oryza sativa.Sci Rep. 2025 Apr 16;15(1):13134. doi: 10.1038/s41598-025-97261-3. Sci Rep. 2025. PMID: 40240775 Free PMC article.
References
-
- van de Velde HJ, et al. NSP-encoded reticulons are neuroendocrine markers of a novel category in human lung cancer diagnosis. Cancer Res. 1994;54:4769–4776. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
