

AhR/miR-23a-3p/PKCα axis contributes to memory deficits in ovariectomized and normal aging female mice
- PMID: 33738140
- PMCID: PMC7940705
- DOI: 10.1016/j.omtn.2021.02.015
AhR/miR-23a-3p/PKCα axis contributes to memory deficits in ovariectomized and normal aging female mice
Abstract
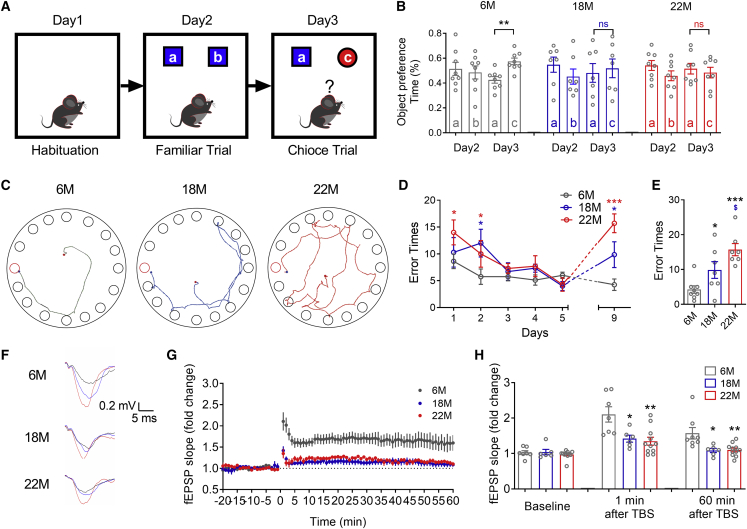
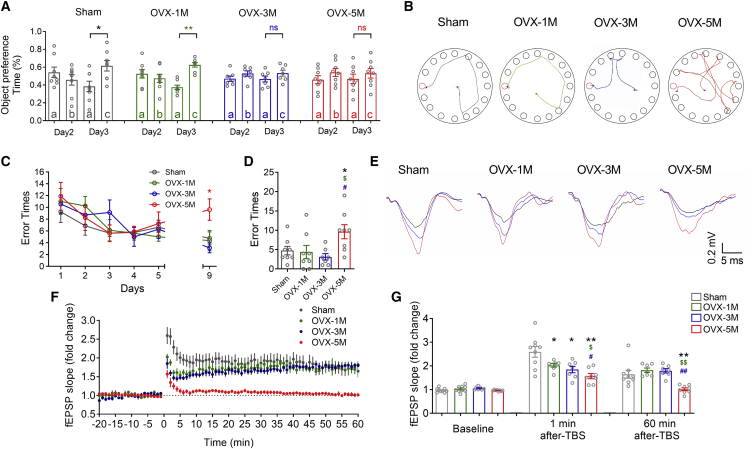
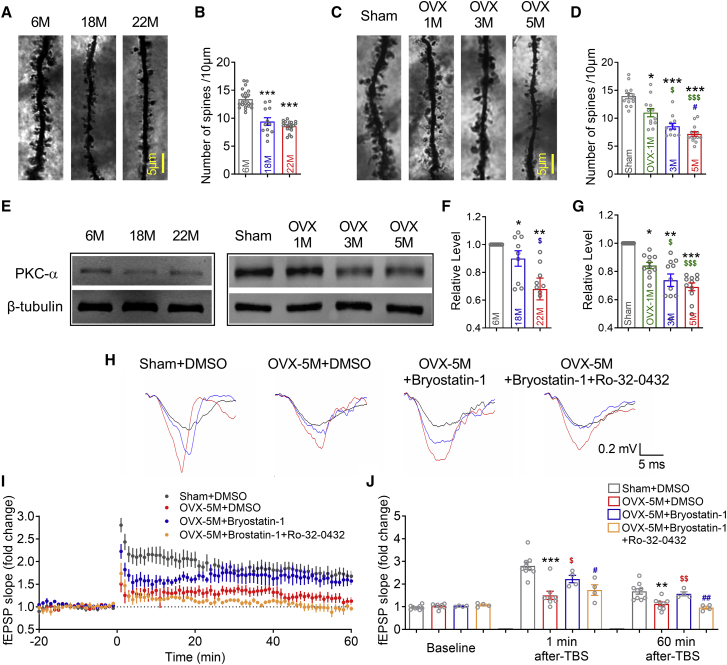
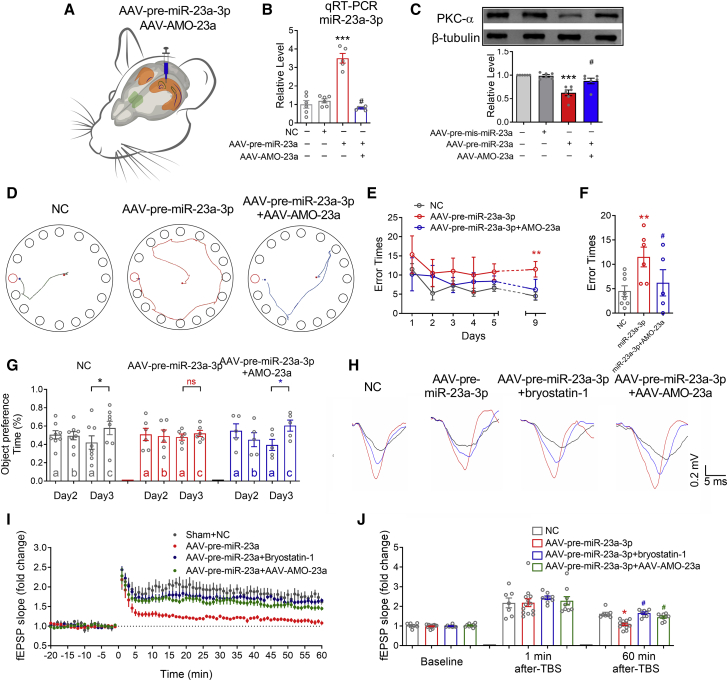
The mechanism of estrogen deficiency-induced cognitive impairment is still not fully elucidated. In this study, we assessed the effect of microRNA (miRNA) on the memory of long-term estrogen-deficient mice after ovariectomy (OVX) and normal aging. We observed that 5-month OVX and 22-month-old normal aging female mice showed significantly impaired spatial and object recognition memory, declined hippocampal long-term potentiation (LTP), and decreased hippocampal protein kinase C α (PKCα) protein. Quantitative real-time PCR analysis showed upregulated miRNA-23a-3p (miR-23a-3p) in the hippocampus of 5-month OVX and 22-month-old female mice. In vitro, overexpression of miR-23a-3p downregulated PKCα by binding the 3¢ UTRs of Prkca mRNAs, which was prevented by its antisense oligonucleotide AMO-23a. In vivo, adeno-associated virus-mediated overexpression of miR-23a-3p (AAV-pre-miR-23a-3p) suppressed hippocampal PKCα and impaired the memory of mice. Chromatin immunoprecipitation analysis showed that aryl hydrocarbon receptor (AhR) binds the promoter region of miR-23a-3p. The AhR-dependent downregulation of PKCα could be prevented by AMO-23a as well. Furthermore, knockdown of miR-23a-3p using AAV-AMO-23a rescued the cognitive and electrophysiological impairments of OVX and normal aging female mice. We conclude that long-term estrogen deficiency impairs cognition and hippocampal LTP by activating the AhR/miR-23a-3p/PKCα axis. The knockdown of miR-23a-3p may be a potentially valuable therapeutic strategy for estrogen deficiency-induced memory deficits.
Keywords: OVX; PKCα; aging; aryl hydrocarbon receptor; memory; miR-23a-3p.
© 2021 The Authors.
Conflict of interest statement
The authors declare no competing interests.
Figures





Similar articles
-
MicroRNA-23a participates in estrogen deficiency induced gap junction remodeling of rats by targeting GJA1.Int J Biol Sci. 2015 Feb 15;11(4):390-403. doi: 10.7150/ijbs.10930. eCollection 2015. Int J Biol Sci. 2015. PMID: 25798059 Free PMC article.
-
MicroRNA-23a mediates mitochondrial compromise in estrogen deficiency-induced concentric remodeling via targeting PGC-1α.J Mol Cell Cardiol. 2014 Oct;75:1-11. doi: 10.1016/j.yjmcc.2014.06.012. Epub 2014 Jun 28. J Mol Cell Cardiol. 2014. PMID: 24984145
-
miR-23a-3p causes cellular senescence by targeting hyaluronan synthase 2: possible implication for skin aging.J Invest Dermatol. 2015 Feb;135(2):369-377. doi: 10.1038/jid.2014.422. Epub 2014 Sep 29. J Invest Dermatol. 2015. PMID: 25264594
-
MicroRNA-23a contributes to hippocampal neuronal injuries and spatial memory impairment in an experimental model of temporal lobe epilepsy.Brain Res Bull. 2019 Oct;152:175-183. doi: 10.1016/j.brainresbull.2019.07.021. Epub 2019 Jul 20. Brain Res Bull. 2019. PMID: 31336125
-
miR-23a-3p/SIX1 regulates glucose uptake and proliferation through GLUT3 in head and neck squamous cell carcinomas.J Cancer. 2020 Feb 10;11(9):2529-2539. doi: 10.7150/jca.30995. eCollection 2020. J Cancer. 2020. PMID: 32201523 Free PMC article.
Cited by
-
Examination of CA1 Hippocampal DNA Methylation as a Mechanism for Closing of Estrogen's Critical Window.Front Aging Neurosci. 2021 Aug 4;13:717032. doi: 10.3389/fnagi.2021.717032. eCollection 2021. Front Aging Neurosci. 2021. PMID: 34421577 Free PMC article.
-
The Causal Role of Magnesium Deficiency in the Neuroinflammation, Pain Hypersensitivity and Memory/Emotional Deficits in Ovariectomized and Aged Female Mice.J Inflamm Res. 2021 Dec 7;14:6633-6656. doi: 10.2147/JIR.S330894. eCollection 2021. J Inflamm Res. 2021. PMID: 34908863 Free PMC article.
-
sVCAM1 in the Hippocampus Contributes to Postoperative Cognitive Dysfunction in Mice by Inducing Microglial Activation Through the VLA-4 Receptor.Mol Neurobiol. 2022 Sep;59(9):5485-5503. doi: 10.1007/s12035-022-02924-1. Epub 2022 Jun 21. Mol Neurobiol. 2022. PMID: 35727436
-
Bazi Bushen capsule attenuates cardiac systolic injury via SIRT3/SOD2 pathway in high-fat diet-fed ovariectomized mice.Heliyon. 2024 May 31;10(11):e32159. doi: 10.1016/j.heliyon.2024.e32159. eCollection 2024 Jun 15. Heliyon. 2024. PMID: 38912487 Free PMC article.
-
Irisin Regulates Microglia M1/M2 Polarization and Promotes Autophagy Through the Sirt3 Pathway to Alleviate POCD.J Neuroimmune Pharmacol. 2025 Apr 11;20(1):34. doi: 10.1007/s11481-025-10201-w. J Neuroimmune Pharmacol. 2025. PMID: 40214830
References
-
- Hamson D.K., Roes M.M., Galea L.A. Sex Hormones and Cognition: Neuroendocrine Influences on Memory and Learning. Compr. Physiol. 2016;6:1295–1337. - PubMed
-
- Yun J., Yeo I.J., Hwang C.J., Choi D.Y., Im H.S., Kim J.Y., Choi W.R., Jung M.H., Han S.B., Hong J.T. Estrogen deficiency exacerbates Aβ-induced memory impairment through enhancement of neuroinflammation, amyloidogenesis and NF-κB activation in ovariectomized mice. Brain Behav. Immun. 2018;73:282–293. - PubMed
-
- Lobo R.A. Hormone-replacement therapy: current thinking. Nat. Rev. Endocrinol. 2017;13:220–231. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
